






Introduction
In fish studies, animal movement information provides fundamental knowledge for understanding fish ecology. Advanced biological information is also important for the construction of fishery management plans (Crossin et al., 2017) and for reducing disease and stress in individuals in aquaculture (Mei et al., 2022). Before technical advances in electronic tags were made, researchers collected information from direct observations at sea by fishing or from mark-recapture data by repeated measures on the same individuals (Hanselman et al., 2014; Lowerre-Barbieri et al., 2019). Since the first tracking of salmon with individual biotelemetric tags, radio-transmitting tags, a global positioning system (GPS), and pop-up satellite archival tags (PSAT) have been developed to follow underwater movements (reviewed in Lowerre-Barbieri et al., 2019).
Tracking fish movement was often called ‘telemetry’ because the technique was based on transmitting data (acoustic or radio signals) from individual tags to the receiver (reviewed in Brownscombe et al., 2019). Acoustic or radio transmitters on animals can provide location signals by communicating with fixed stations or mobile receivers (see Hussey et al., 2015). Recently, ‘bio-logging’ has been widely used for data logging and transmission of animal movement research across different taxa (Chung et al., 2021), including in fish (Lowerre-Barbieri et al., 2019). Hereafter, the term ‘bio-logging’ is used throughout this study to indicate fish-tracking studies using devices.
Yellowtail (Seriola quinqueradiata) is a temperate migratory fish in the Northwest Pacific Ocean (Dong et al., 2020; Jeong et al., 2016). Previous studies from multiple sampling of eggs and juveniles revealed that they are distributed from the East China Sea continental shelf to the Pacific coast of Japan (Tian et al., 2012) and is known to show large migratory routes for feeding and south for spawning (Sassa et al., 2020). In East Asia, it is one of the most important species for aquaculture species of its high edible and commercial value (Sicuro & Luzzana, 2016) but the amount of yellowtail catches is recently changing (Tian et al., 2012) and the detailed GPS tracking studies have been limited. Therefore, it is required to collect data on monitoring of continuous movement and seasonal distribution of the species. In this study, we aimed to investigate the movement pattern of yellowtail inhabiting the East Sea from October to December. We confirmed the relationship between the migration route of the yellowtail and sea surface temperature. Additionally, we examined how the month and solar elevation influence their swimming depth.
Materials and Methods
On 14th October 2022, we deployed PSATs (miniPAT, Wildlife Computers, Redmond, WA, USA), 118 mm (length) and 38 mm (diameter), 61 g (weight in air) on two yellowtails weighing 8.7 kg and 9.5 kg (Fig. 1) in South Sea of Korea (latitude: 35°0'4'' N, longitude: 129°10'19'' E). The fish were purchased from a commercial fishery (Yookilsusan, Tongyeong, Korea) on March 19 2020, and were kept before deployment in a sea cage (6 m × 12 m × 6 m) at Tongyeong Megacosm Test Station operated by the Korea Institute of Ocean Science and Technology. This PSAT tag provides depth (range: 0–1,700 m, resolution: 0.5 m, accuracy: ± 1% of reading) and temperature data (range: –40°C to 60°C, resolution: 0.05°C, accuracy: ± 0.1°C) with light sensor for geolocation (for a detailed spec, see Wildlife Computers Webpage, https://wildlifecomputers.com/our-tags/pop-up-satellite-tags-fish/minipat/). For testing the PSAT deployment, captive yellowtails were previously tested and confirmed with physiological measurements for tagging (Oh & Jeong, 2021). We set the miniPAT device to collect depth and temperature data at 5-minute intervals. The daily sunset and sunrise data are recorded based on the calculations from the collected light data. Also, the mixed layer temperature, depth and temperature profile, and histograms are summarized and recorded every 12 hours. After the release of the fish with the attached device, the equipment will be activated when the fish reaches a depth of 5 meters or deeper. The device has a pre-set automatic release function that causes it to float to the surface after 90 days from the time it was attached to the fish. When the device floats to the surface, it uploads the collected data through ARGOS satellites.
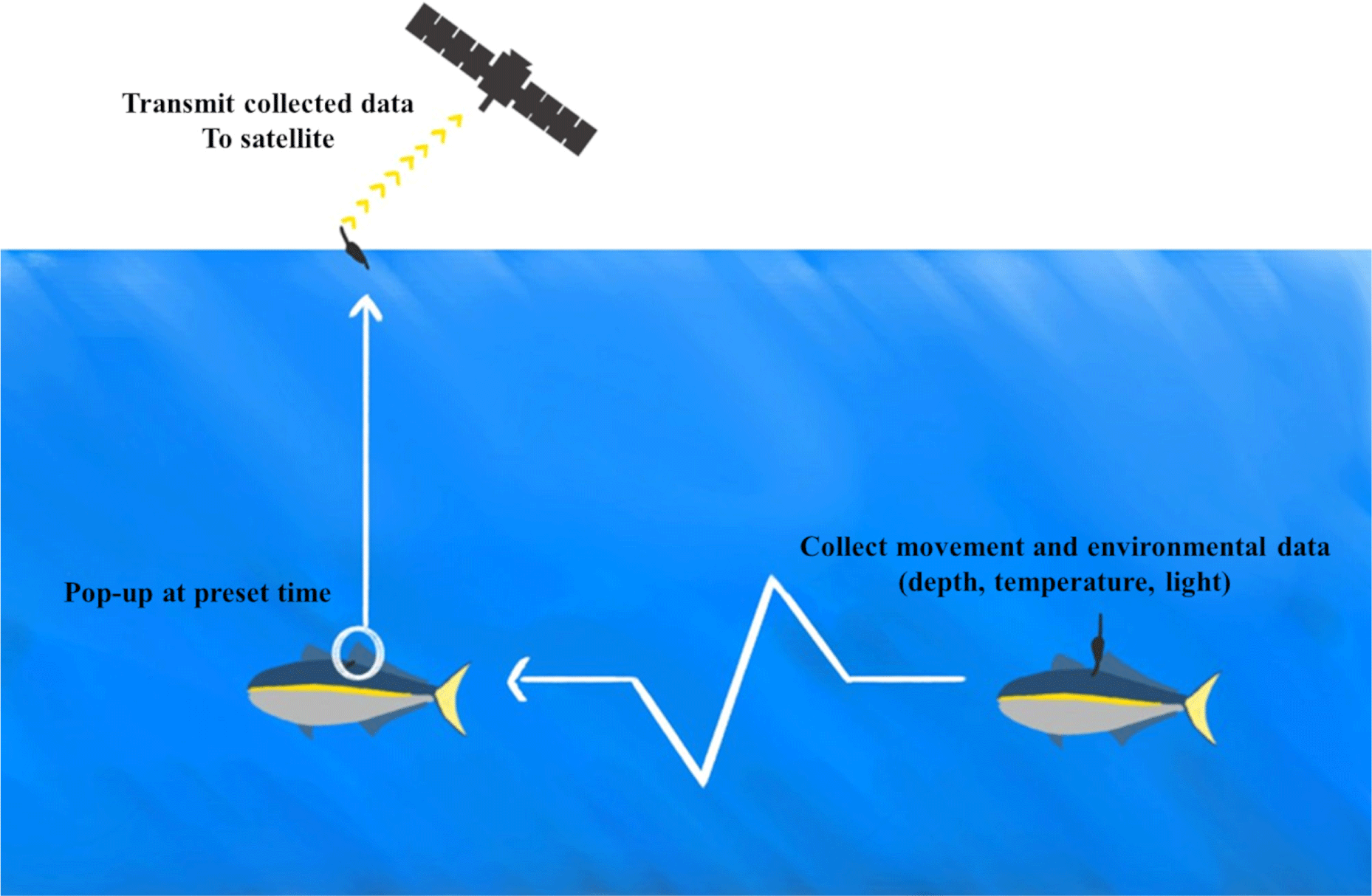
We estimated the daily location of each fish using Wildlife computers GPE3, which is based on the hidden Markov model methods of Pedersen et al. (2011) and Basson et al. (2016). The GPE3, global position estimator was developed by the manufacturer of the miniPAT device we used in our research. It uses observational data such as sunrise and sunset data calculated from collected light data, as well as sea surface temperature and maximum dive depth data, to estimate the fish’s daily location (Hill & Braun, 2001; Wildlife Computers, 2012). The estimator also utilizes a random walk movement model. In addition, we inputted the device deployment date and location, as well as the first ARGOS location data to identify the pop-up date and location. Based on a previous yellowtail bio-logging study (Kim et al., 2021), we set a typical traveling speed of 1.0 m/s for yellowtail fish.
The collected depth and temperature data with depths of 1.5 meters or shallower were excluded from the analysis. We calculated the daily average depth and temperature based on UTC + 9 time zone. values for depth and temperature. Then, based on daily data, we calculated the overall average depth and temperature, along with the monthly averages for temperature and depth. Therefore, we used the ‘solrad’ package (Seyednasrollah, 2018) in R 4.2.0 and interpolated location data about yellowtail to calculate solar altitude at each recorded time with dive depth data. We defined time periods when the solar elevation was greater than 0 as “daytime” and the time periods when it was less than or equal to 0 as “nighttime”.
To understand the movement of yellowtail, we considered environmental factors such as sea surface temperatures, length of daytime, and length of nighttime. The sea surface temperatures of the East Sea were obtained from the MODIS of the Aqua satellite. We used the monthly ocean color map of SSTs for the period from October 2020 to December 2020 in the geographical range of 32°N–42°N and 124°E– 133°E.
Results
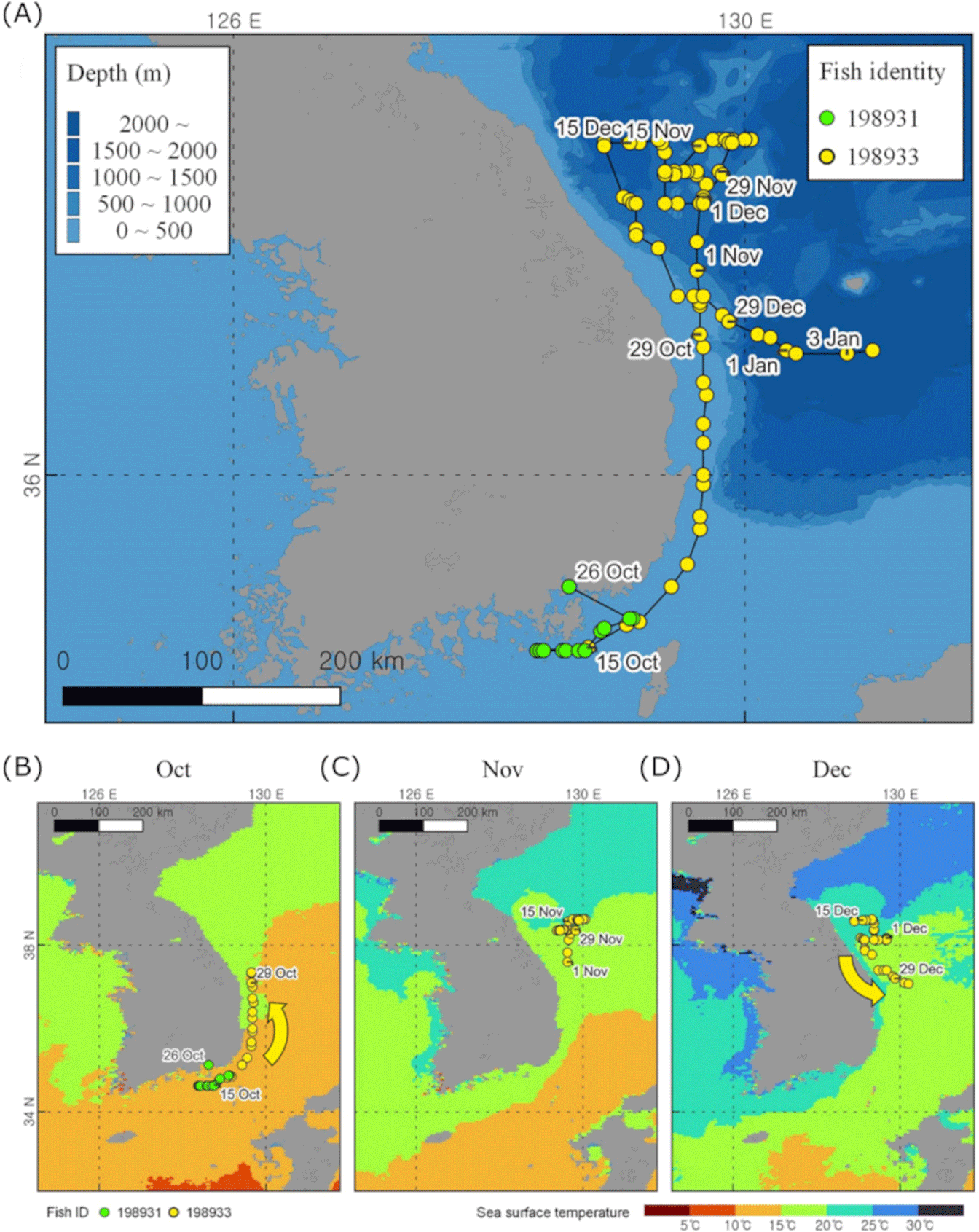
For the two yellowtails, we acquired 12 days of recording from ID198931 from 14 October to 26 October and 82 days of continuous recording from ID198933 from 14 October to 4 January (Fig. 2A). The average swimming depth for ID198931 and ID198933 were 8.20 ± 2.56 m and 24.95 ± 9.33 m, respectively, and the average ambient temperature for both was 20.65 ± 0.36 and 16.52 ± 1.86 degrees Celsius, respectively. ID198931 traveled an average of 15.3 km per day and for 12 days, traveled 176.2 km. ID198933 traveled an average of 14.7 km per day and a total of 1172.4 km over a period of 82 days (Table 1). We examined the monthly tracking data and found that ID 198933 fish migrated northward to latitude 38°37'30'' in October when the ocean surface temperature was warm (Fig. 2B and 2C). In December, during the period of the cold sea surface that sea surface, the fish moved southward (Fig. 2D). The collected temperature data exhibited a similar pattern, showing lower temperatures in November and December compared to October (Fig. 3B).
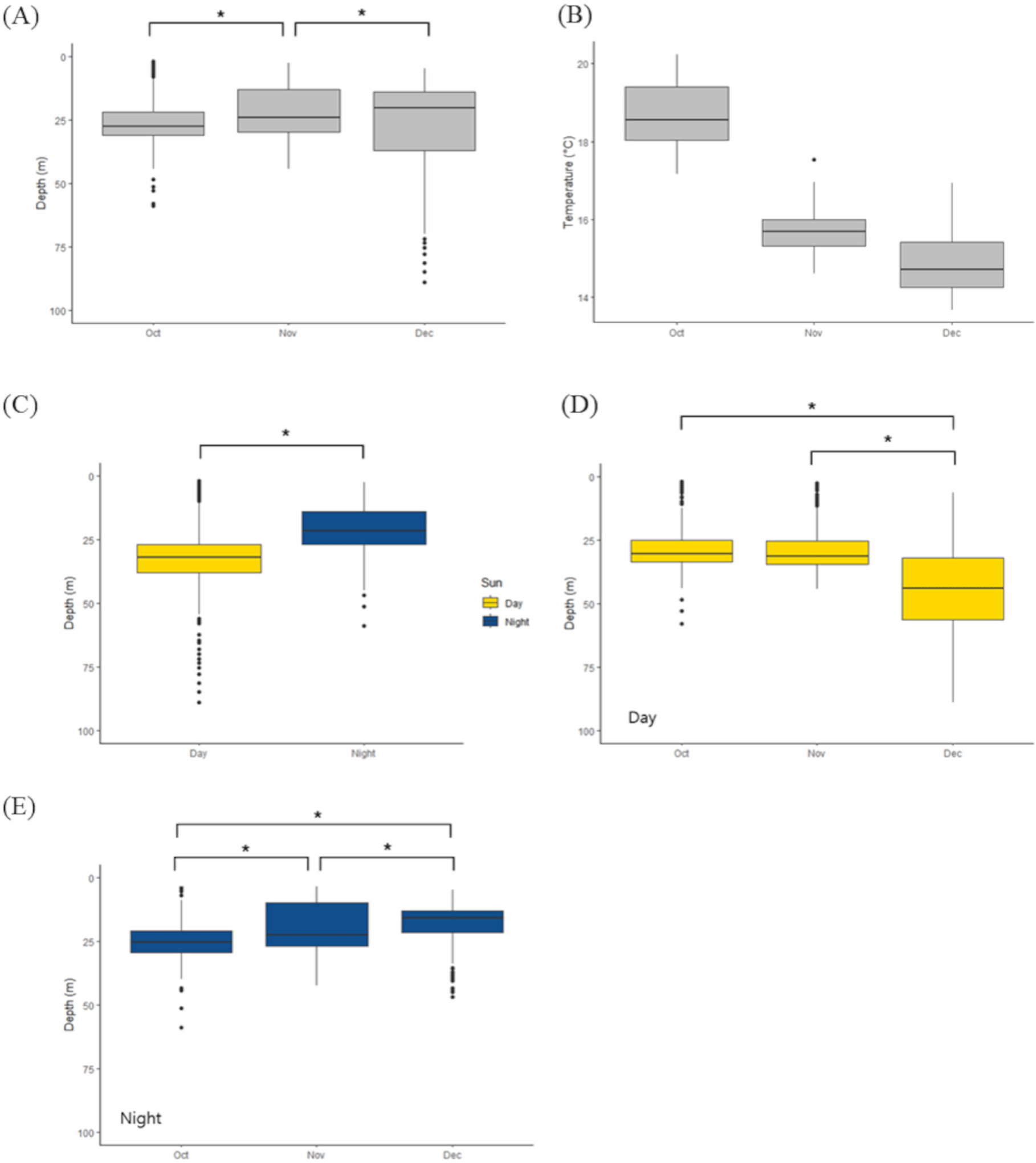
To estimate the effects of time and seasonal change in swimming depth in ID 198933, we conducted a general linear model (explanatory variable: month [October or November or December] and time [day or night], and responsible variable: depth) with Tukey post-hoc test. We found that the average depth varies every month. In November, the average swimming depth was 22.28 ± 9.92 m, however October (26.47 ± 8.34 m) and December (27.31 ± 17.53 m) showed deeper depth (Fig. 3A). Also we found that swimming depth in the daytime (33.70 ± 14.80 m) is deeper than those in nighttime (20.65 ± 8.44 m; between day and night, p < 0.001, z = –42.42, Fig. 3C). Then we conducted a general linear model only with daytime depth (explanatory variable: month [October or November or December], and responsible variable: daytime depth) or nighttime depth (explanatory variable: month [October or November or December], and responsible variable: nighttime depth). With this separated model, we found that daytime swimming depth in December (43.74 ± 18.16 m) is deeper than those in October (29.04 ± 9.06 m) and November (28.24 ± 9.52 m) (between October and December, p < 0.001, z = –19.470; between November and December, p < 0.001, z = –21.383, Fig. 3D) and nighttime swimming depth became shallower from October (25.02 ± 7.53 m) to December (November 19.70 ± 8.93 m, December 17.73 ± 6.71 m; between October and November, p < 0.001, z = 15.501 November and December, p < 0.001, z = 6.748; between October and December, p < 0.001, z = 20.082, Fig. 3E).
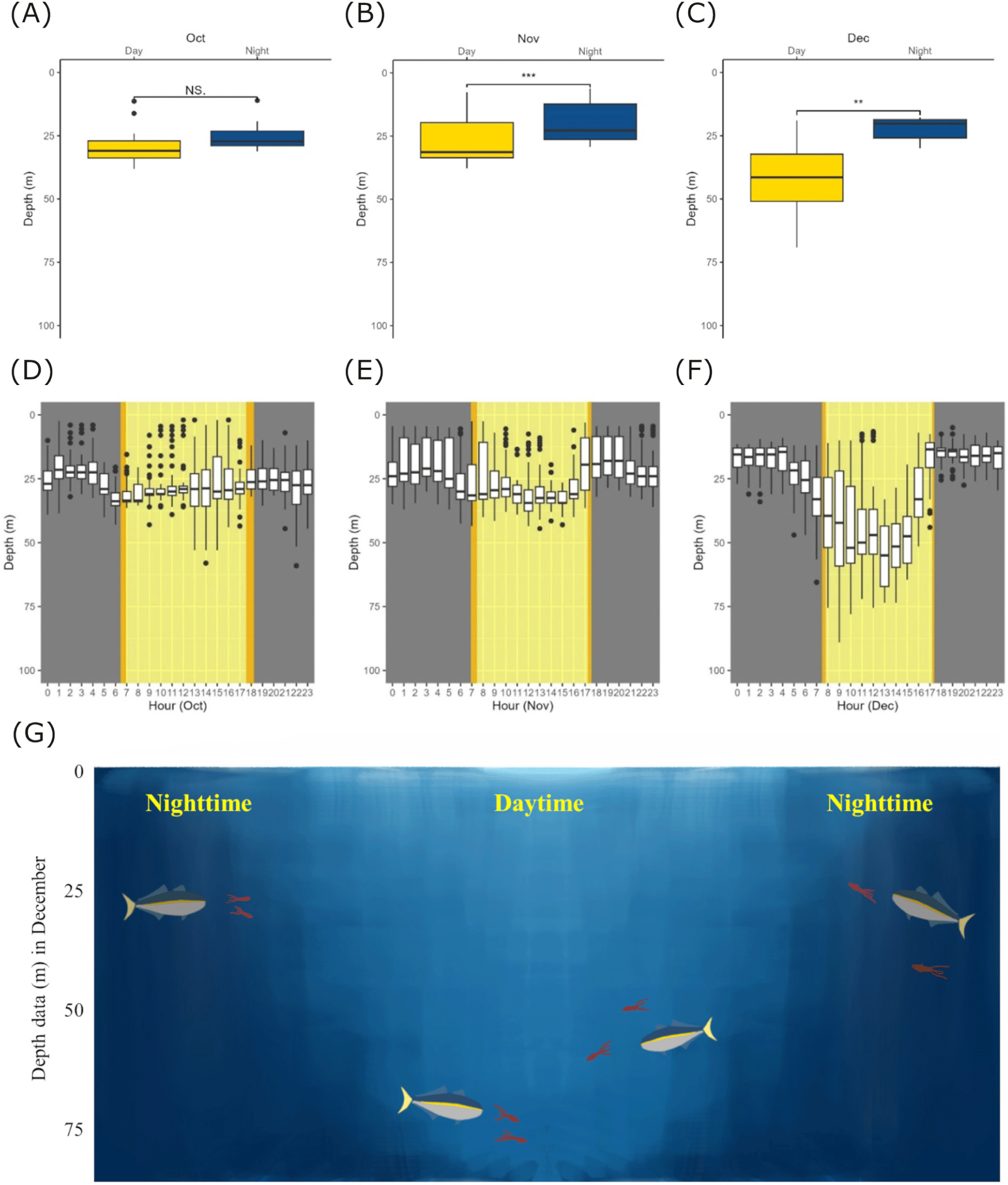
From the long-monitored individual (ID198933), we further investigated its monthly changes and diel swimming behavior (Fig. 4). We compared the average daily depth values for daytime and nighttime within each month (October, November, and December) using paired t-tests (Fig. 4A, 3C, and 3D). Compared to October, the swimming depths during both daytime and nighttime showed significant differences in November and December. In November, the average swimming depth during daytime was 27.07 ± 9.16 m (average ± SD) and the average swimming depth during nighttime was 20.10 ± 8.05 m (average ± SD; between daytime and nighttime in October, t = 4.8872, p = 0.0003) and in December, mean depth in daytime was 42.42 ± 15.29 m (average ± SD) and mean depth in nighttime was 22.30 ± 4.84 m (average ± SD; between daytime and nighttime in December, t = 3.6037, p = 0.0069). Furthermore, we divided daily depth data for October, November, and December into hourly intervals (24 hours) and represented them (Fig. 4D, 4E, and 4F). This boxplot shows deeper swimming depth during daytime compared with nighttime in November and December.
Discussion
According to our research, released yellowtail moves northward during October and mid-November. At latitude around 38.6 degrees, it stopped moving northward and exhibited a slow swimming speed and then they moved south from mid-December. We thought the movement track of the ID 198933 fish is affected by sea surface temperature (Fig. 2) and also warm currents in the East Sea (Fig. 5). Additionally, we investigated how swimming depth differed during day and night. A yellowtail was distributed deeper during the daytime (Fig. 3C), and their swimming depths were deeper in winter (Fig. 3D). Furthermore, there are significant differences in swimming depths between day and night which were observed in November and December (Fig. 4). In this research, we consider to understand horizontal and vertical distribution of yellowtail in the East Sea through the analysis of their tracking and depth data.
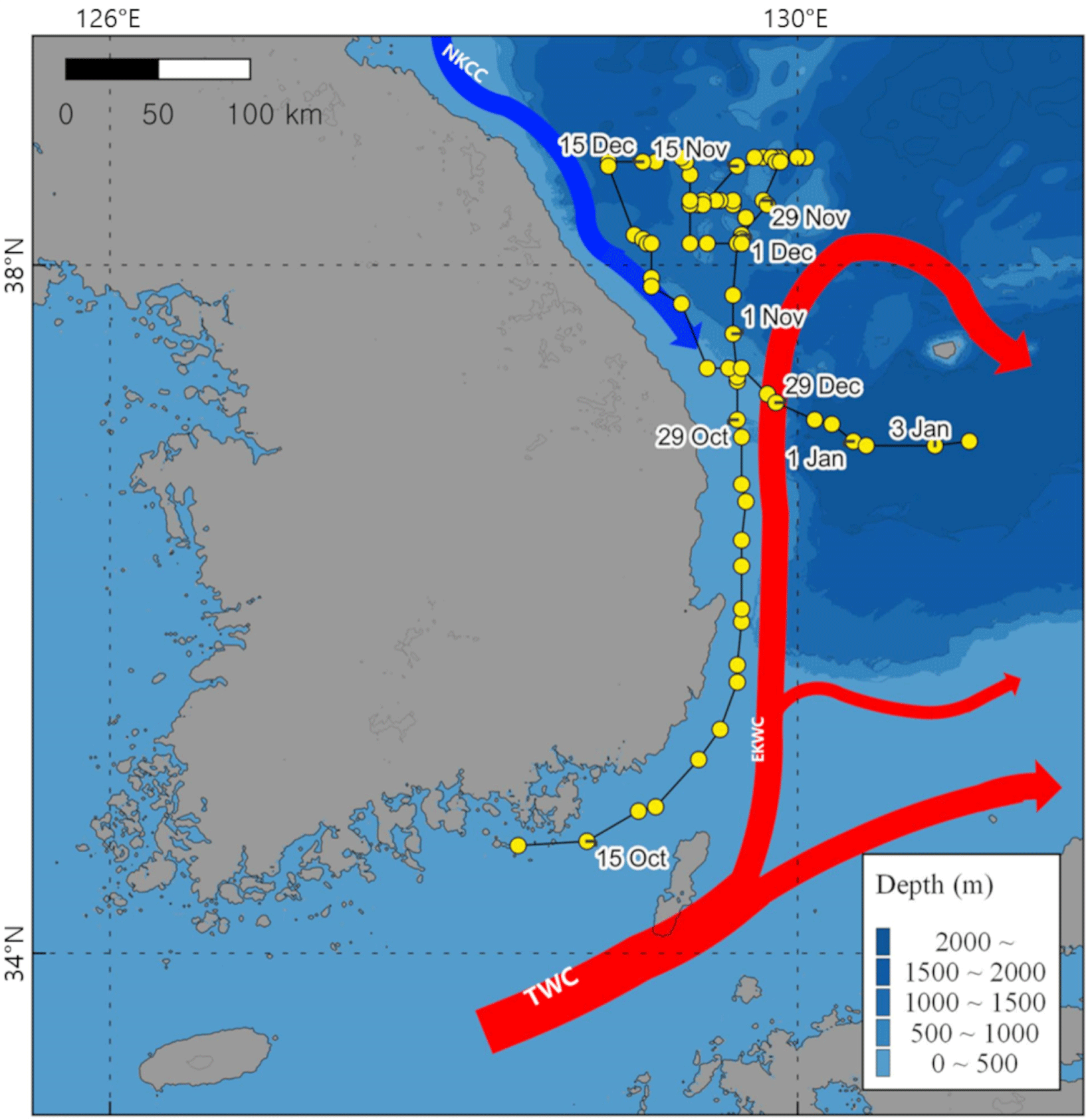
The distribution of the yellowtail is highly correlated with the surface temperature of the ocean (Tian et al., 2012). According to a previous yellowtail tracking study, they migrated north for feeding during the Summer and Autumn when the surface of the ocean temperature is warm (Furukawa et al., 2020; Ino et al., 2008), however, during the winter when water temperature decreases, there is a significant migration of yellowtail towards southward spawning grounds in the East China Sea and West Kyushu regions (Ino et al., 2008; Sassa et al., 2020). Furthermore, the fact that yellowtail is associated with sea surface temperature demonstrates its close relationship with warm currents. Several Japanese research teams suggest that the main heat source in the East Sea, known as the Tsushima Warm Current (TWC), plays a crucial role in the migration route of the yellowtail (Tian et al., 2008, 2012). Additionally, the flow direction of the East Korean Warm Current (EKWC), which is the main branch of the TWC, overlaps with the October migration route of yellowtail in our research (Fig. 5). These results demonstrate that the migration and distribution of yellowtail on the East side of the Korean peninsula are related to sea surface temperatures and the flow of warm currents, EKWC.
There are many studies on the migration pattern of predator species related to the distribution pattern of prey species (Furey et al., 2018). The seasonal distribution of yellowtails is also affected by their prey species. In the East Sea, there are many pelagic small species such as Japanese common squid (Todarodes pacificus), Japanese anchovy (Engraulis japonicus), and horse mackerel (Trachurus japonicas) which are the main prey for yellowtail (Tian et al., 2008). Particularly, the Japanese common squid prefers the East Sea region from June to November (Alabia et al., 2016) and their monthly distribution patterns have a correlation with the monthly variations of the northern boundary of the TWC (Choi et al., 2008). Japanese anchovies are also sensitive to changes in oceanographic conditions such as ocean temperature (Jung et al., 2014), and exhibit different distribution patterns depending on the season (Bang et al., 2022). These seasonal distribution changes of the prey species could affect habitat selection and migration behavior of yellowtails, and future research on these of the East Sea of Korea is required.
In our research, yellowtails exhibited different swimming depths during day and night (Fig. 4), which can be explained by following prey species that display diel vertical migration. Zooplankton is the most prominent example which shows diel vertical migration patterns (Hays, 2003). It results in predator avoidance during the daytime in deep water and feeding behavior during the night in shallow surface water (van Haren & Compton, 2013). Recent studies have also revealed diel vertical migration in various fish and shark species (Andrzejaczek et al., 2021; Dypvik et al., 2012a). Lanternfish (Benthosema glaciale), a key species in polar ecosystems, exhibit diel migration patterns depending on the movement of seasonally migrating zooplankton (Dypvik et al., 2012b). Whale sharks, the giant plankton hunter, also have a diel vertical migration pattern, and it might be influenced by the distribution of their zooplankton prey (Andrzejaczek et al., 2021). Furthermore, diel vertical migration has been observed in various pelagic fish and shark species, such as the bigeye thresher shark (Coelho et al., 2015; Preti et al., 2008) and blue shark (Stevens et al., 2010). Especially, various small pelagic species including common squid, the main prey source of yellowtail, displayed a vertical migration pattern (Kawabata et al., 2006) which could lead to frequent short dives of yellowtail through the deep thermocline layer (Furukawa et al., 2020).
A previous study was reported to track a yellowtail near Jeju Island in the South Sea of Korea for 40 days in December and January (Kim et al., 2021). It also shows that the yellowtail is diving deeper during the day. This suggests that yellowtails would have diel swimming behavior, at least in winter. Although the tracking area did not overlap with ours and the period was relatively short, the earlier study near Jeju Island presented deeper (40–70 m depth and maximally up to 170 m in Kim et al. [2021]; 10–40 m depth and maximally 89 m in our study). Such a difference could be related to their migration swimming. We had 1,170 km of total traveling to the north along the East Sea coast for migration while the earlier study individual remained near Jeju Island (Kim et al., 2021). Thus, we suspect that our yellowtail had a migratory swimming during winter and it was reflected to be shallower depths for the tracking period.
Recent climate change could have significant impacts on the marine ecosystem of the East Sea (Lenoir et al., 2020). The effects of climate change on sea surface temperature have been widely observed across numerous marine regions (Belkin, 2009). In particular, the East Sea exhibited a consistent warming trend from 1982 to 2018 (Lee & Park, 2019). As previously discussed, the distribution of various marine species inhabiting the East Sea is related to sea surface temperature (Alabia et al., 2016; Jung et al., 2014). Therefore, the warming trend in ocean temperature may influence the distribution of these species. Especially, the distribution of yellowtail, a predatory pelagic fish, is affected by annual sea surface temperature variations (Lee & Go, 2006; Tian et al., 2012). Bio-logging studies on marine species are essential to demonstrate how various fish species respond to ocean warming trends (Lowerre-Barbieri et al., 2019). That is why the study of fish movement and distribution using bio-logging devices will become increasingly important.
The present experiment indicated that the horizontal migratory route of yellowtail was in accordance with the route of East Korea Warm Current which is the main branch of Tsushima Warm Current in the fall and early winter seasons, and showed significant diel vertical movement patterns from November and December. This information may be helpful to biologists who study geo-ecological knowledge and are interested in directions for future research of marine fish’s relation to prey and climate change.