
Introduction
Cryopreservation is a valuable technique that assists in the genetic improvement of cultured gametes and provides a continuous supply of high-quality sperm for artificial insemination. It is of interest not only for fish farming but also for the conservation and genetic improvement of resources. Cryopreservation has been successfully established in freshwater fish species, such as salmons, sturgeons, and carps, and has recently been actively researched in marine fish (Le et al., 2011a, 2011b). Fish sperm preservation is advantageous for solving the unbalanced sex ratio and the asynchronous eggs and sperm discharge time between females and males during spawning season (Chang et al., 1997). Thus, sperm cryopreservation at the gamete level may have many advantages and is already being used in broodstock and medical fields worldwide. Due to the lack of awareness of cryopreservation in Korea thus far, it is necessary to develop basic technologies that can be applied to the actual aquaculture field and conduct physiological research on cryopreserved sperm in aquaculture. In addition, because the types and concentrations of cryoprotective agents (CPAs) and diluents can cause remarkable differences in cryopreservation results, basic information on cryopreservation of fish sperm is necessary (Lee et al., 2021). CPAs play a role in alleviating and regulating conditions unfavorable to cell survival, such as concentration of intracellular electrolytes, increase in osmolality, and formation of ice crystals inside and outside cells, which are generated during the freezing of cells (Ahn et al., 2018). The diluted solution actively helps in the survival of the sperm after freezing and thawing and prevents the sperm from being activated by osmotic shock, with osmolality similar to that of the semen used (Kho & Kang, 2003; Zidni et al., 2020). Present studies on cryopreservation of the family Sciaenidae have been conducted by many researchers, both domestic and foreign (Gwo, 1993; Gwo, 1994; Le et al., 2011a; Lim et al., 2010). The first protocol for large yellow croaker sperm cryopreservation was published using different CPAs and diluents (Lee & You, 2002; Lin et al., 2006; Xiao et al., 2007; Xu et al., 2014). However, effective storage methods for gametes of large yellow croaker (Larimichthys crocea) have not been identified.
The study species, large yellow croaker, belonging to the family Sciaenidae, is an economically important fishery resource in Korea. Currently, the quantity of domestic resources has decreased significantly, and the dependence on imports is gradually increasing. Seed production of large yellow croaker faces many problems as follows: 1) the sensitive nature of the fish results in severe stress during transportation; 2) many cases of death due to injuries caused by severe struggle have been reported; 3) when seeds are produced, the sex ratio is not balanced or the timing of the male and female spawning and spinning is not synchronized. Thus, to solve these problems, it is necessary to restore the resources through aquaculture. Accordingly, this study aimed to develop a cryopreservation protocol for large yellow croaker.
Materials and Methods
The experiment was conducted from April to July 2020. Adult large yellow croakers were reared in a fiber cylinder fish tank equipped with recirculation systems containing seawater, at a temperature range of 16°C–19°C, with salinity of 30.2 ± 1.2 psu, dissolved oxygen of 7.1 mg∙L−1, and pH of 8.1, at Ocean & Fisheries Science Institute Resources Creation Research Institute, Shinan-gun, Korea. The mean length and body weight of males (n = 15) were 19.4 ± 2.7 cm and 80.67 ± 4.53 g, respectively. During the study, the fish were fed with a commercial diet once a day in the morning. After acclimation, fish were anaesthetized with tricaine methanesulfonate (MS-222) and Ovaprim (0.1 mL kg−1; Syndel Laboratories, Nanaimo, BC, Canada) was injected into the ventral muscle (intraperitoneally) of the males. To prevent any mixing with feces, blood, water, urine, and mucous, the area around the genitals of male fish was cleaned using paper towels. The sperm was stripped into plastic syringes (1.0 mL) by applying gentle pressure to the ventral muscle and the syringes containing sperm were then placed in an icebox and directly taken to the laboratory for analysis, observation, and cryopreservation. The sperm samples selected for use in the study had a total motility of more than 80% and a sperm activity index (SAI) of 5.
Sperm motility was analyzed after collecting the sperm samples. To measure sperm motility, sperm samples were diluted in seawater at a ratio of 1:100 (10 µL fresh sperm to 1,000 µL seawater). Subsequently, 10 μL of the mixture (sperm + sea water) was dropped onto a glass slide without a cover slip and observed under a microscope at 200× magnification (CH30, Olympus, Tokyo, Japan). To investigate the post-thawing sperm quality after each cryopreservation treatment, movable sperm ratio (MSR) and SAI were estimated. MSR was calculated according to the method described by Ahn et al. (2018), which states that MSR is the ratio between moving sperm count to the total count of fresh/post thawing sperm investigated with a microscope at 200× magnification (CH30, Olympus). SAI was measured as follows: (index score × motile sperm ratio [%]) / 100. The calculation of the index score (scoring method) was based on the characteristics of the movement of the sperm (quick, slow, vibration, and motility) (Table 1). To evaluate MSR and SAI of cryopreserved sperm, artificial seawater containing 27 g NaCl, 0.5 g KCl, 1.2 g CaCl2, 4.6 g MgCl2, and 0.5 g NaHCO3 per liter of distilled water was used as a diluent at a ratio of 1:99 (sperm: artificial seawater) according to the protocol by Le et al. (2011b). Subsequently, 1 µL of the mixture was placed onto a glass slide (Teflon printed glass slide; 21 wells; diameter of each well, 4 mm; Funakoshi, Tokyo, Japan) without a cover slip. Furthermore, cryopreserved sperm quality was investigated immediately after thawing in a 20°C water bath for 10 s through observation under a microscope at 200× magnification. Triplicate experiments were conducted for each treatment and replicated three times.
| Index | Score | Motility characteristics |
|---|---|---|
| I | 5 | Rapid forward movement |
| II | 4 | Slow forward movement |
| III | 3 | Slow forward movement and moderate vibrating movement |
| IV | 2 | Slow vibrating movement |
| V | 1 | Immobility |
In the first experiment, five cryoprotectants, ethylene glycol (EG), dimethyl sulfoxide (DMSO), propylene glycol (PG), glycerol, and methanol, were tested for their effect on sperm quality after cryopreservation in large yellow croaker. All cryoprotectants were applied at a final concentration of 10%. Cortland solution was used as a diluent, and each mixture (CPA + diluent) was placed into a 1.5 mL tube. The ratio between sperm and the mixture was 1:3. At this stage, the equilibration time was not adjusted. The mixture was applied to 0.25 mL plastic cryopreservation straws that were subsequently sealed and frozen. The sperm was first frozen for 2 min in a tray (size: 8.5 cm wide and 20 cm long) at 3 cm above liquid nitrogen (LN) in a Styrofoam box, with a lid covering the box to maintain the first freezing temperature; then, it was submerged in LN (−196°C) for the second phase of freezing (Ahn et al., 2018; Le et al., 2008; Lim et al., 2007). Fresh sperm from the same male fish was considered the control.
The second experiment was conducted to observe the effects of different diluents, including Stein’s solution (SS), Hanks’ balanced salt solution (HBBS), marine fish Ringer’s solution (MFRS), artificial seminal plasma (ASP) of small yellow croaker, and Cortland solution, on sperm preservation. The contents of the diluents are presented in Table 2. The composition of ASP was based on the biochemical properties of small yellow croaker seminal plasma (Lim et al., 2010). The CPAs and diluents were precooled at 4°C before use (Lee et al., 2021; Zidni et al., 2020). Sperm was separately mixed with various diluents at 10% EG. Each mixture was frozen and thawed using the same technique as described above.
| Ingredient |
Stein’s solution (Stein & Bayrle, 1978) |
Hanks’ balanced salt solution (HBBS) (Bart et al., 1998) |
Marine fish Ringer’s solution (MFRS) (Le et al., 2011) |
Small yellow croaker ASP (Le et al., 2011) |
Cortland solution (Cheng et al., 2015) |
|---|---|---|---|---|---|
|
NaCl (Sodium chloride, g/L) |
0.75 | 8.00 | 13.50 | 9.92 | 7.25 |
|
KCl (Potassium chloride, g/L) |
0.038 | 0.40 | 0.60 | 0.77 | 0.38 |
|
CaCl2 (Calcium chloride, g/L) |
- | 0.14 | 0.35 | 0.13 | 0.18 |
|
MgSO4 · 7H2O (Magnesium sulphate, g/L) |
- | 0.20 | - | - | 1.00 |
| Na2HPO4 · 7H2O (g/L) | - | 0.06 | - | - | 0.23 |
|
KH2PO4 (Potassium phosphate, g/L) |
- | 0.03 | - | - | 0.41 |
| Hen egg yolk (mL) | 100 | - | - | - | - |
|
C6H12O6 (Glucose, g/L) |
0.10 | 1.00 | - | 1.00 | 1.00 |
|
NaHCO3 (Sodium bicarbonate, g/L) |
0.20 | 0.17 | 0.03 | - | - |
|
MgCl2 (Magnesium chloride, g/L) |
- | - | 0.02 | 0.05 | - |
|
NaH2PO4 · 2H2O (Sodium phosphate, g/L) |
- | - | - | - | 1.00 |
To measure the optimum EG concentration, a third experiment was conducted because EG was found to be the most efficient CPA for cryopreservation in the first experiment. Different concentrations of EG (5%, 10%, 15%, and 20%), were tested with ASP as the diluent. The process of freezing and thawing was the same as that in the previous experiments.
The prior experiments demonstrated that a combination of 10% EG and ASP were optimum for sperm cryopreservation in large yellow croakers. To investigate the effect of the dilution ratio, the sperm was diluted at ratios of 1:1, 1:3, 1:5, 1:10, 1:30, 1:100, and 1:200 with ASP containing EG at a final concentration of 10%. The time of equilibration was not specifically set. The freezing and thawing method was the same as described in the prior experiments.
To investigate the optimal equilibrium time, the sperm was diluted at a 1:1 ratio with ASP containing EG at a final concentration of 10%, and the mixture was then allowed to equilibrate for various lengths of time (30, 60, 90, 120, 150, 180, 210, 240, 270, and 300 s). During equilibrium, sperm samples were loaded into 0.25 mL straws (3 replicate straws × 10 equilibrium times) and sealed with sealing powder (Reproduction provisions, LLC, Walworth, WI, USA) at room temperature 22-24.5 °C.
To investigate the optimal freezing rate, the sperm was frozen under four different freezing rates (–1°C, –5°C, –10°C, and –20°C/min) using a CRF (14S, SY-LAB, Neupurkersdorf, Austria). The controlled rate freezer was set to start from 10°C to –80°C for the first freezing, and then the frozen sperm was stored in LN. During the thawing method, the straws were removed from the LN storage using tweezers and immediately immersed for 10 s in a water bath at different temperatures (20°C, 25°C, 30°C, 35°C, 40°C, 45°C, and 50°C). The straws were removed with tweezers and both sealed ends of the straws were cut off using a straw cutter. Thawed sperm were stored in a microtube and used for evaluation of post-thaw sperm motility.
Even when using the optimal CPA and diluent selected from the previous experiment, the sperm suffered DNA damage during the cryopreservation and thawing processess. SCGE assay (Comet Assay, Trevigen, Gaithersburg, MD, USA) was performed to evaluate DNA damage, and fluorescent staining using SYBR Gold (dilution × 10,000 DMSO) was performed for 30 min, followed by observation under a fluorescence microscope (DM 2500 microscope, Leica, Wetzlar, Germany). Head intensity (DNAh), tail intensity, and tail migration were assessed using Comet Assay IV lite System (Instem, Staffordshire, UK). The percentage of tail DNA was calculated using the following equation: [100 × (DNAc – DNAh)/DNAc] (Cabrita et al., 2005; Lee et al., 2021; Zidni et al., 2020), where DNAc and DNAh are the sum of the intensities of the pixels in the whole comet area and the head area, respectively (Pérez-Cerezales et al., 2009). The degree of damage was categorized and the percentage of cells within each degree of damage was established.
Data were expressed as the mean ± SE. Statistical evaluation was performed using one-way analysis of variance with IBM SPSS Statistics 25.0 (USA). Means were analyzed using Duncan’s multiple range test, and differences were considered significant at p < 0.05. The relationships between sperm motility, survival rate, and DNA damage were tested using bivariate Pearson correlation.
Results
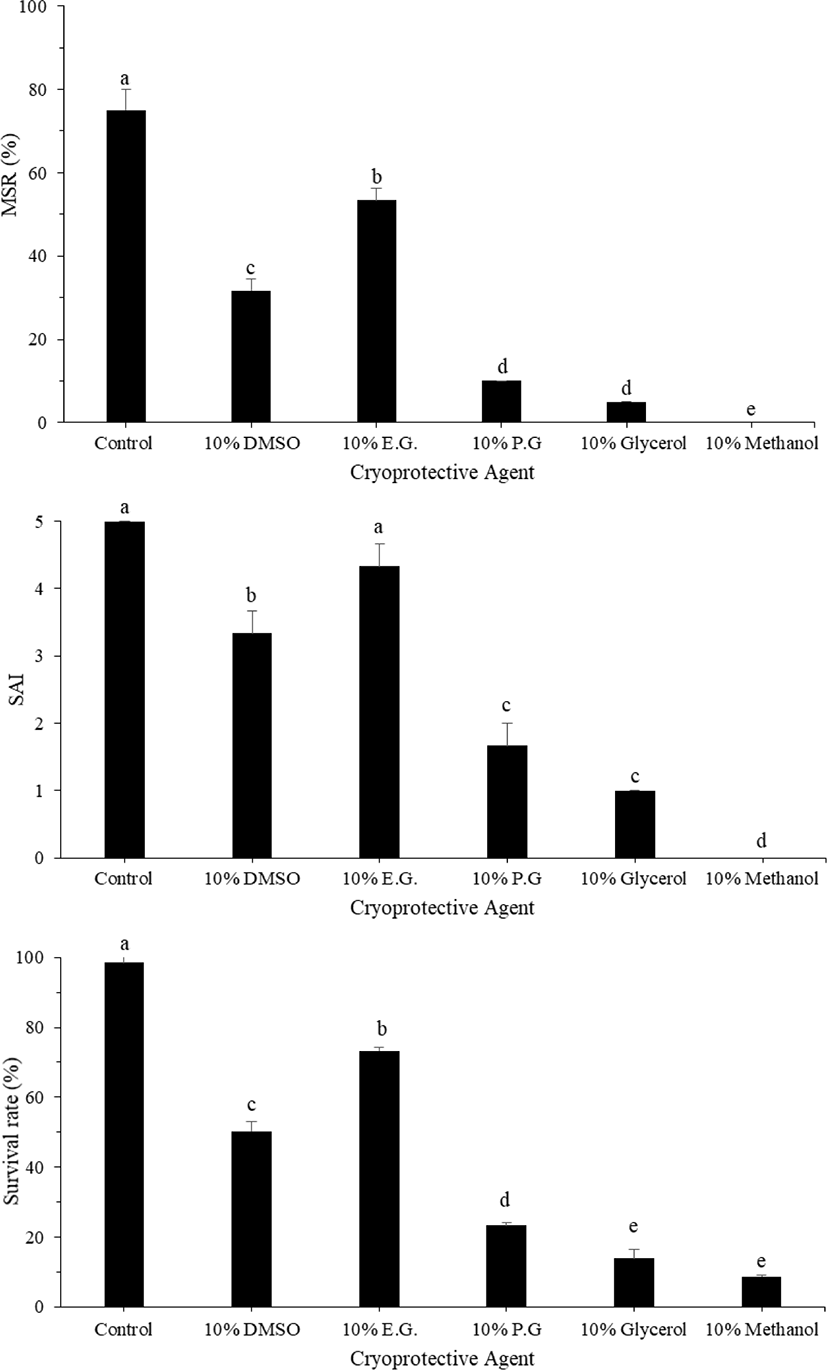
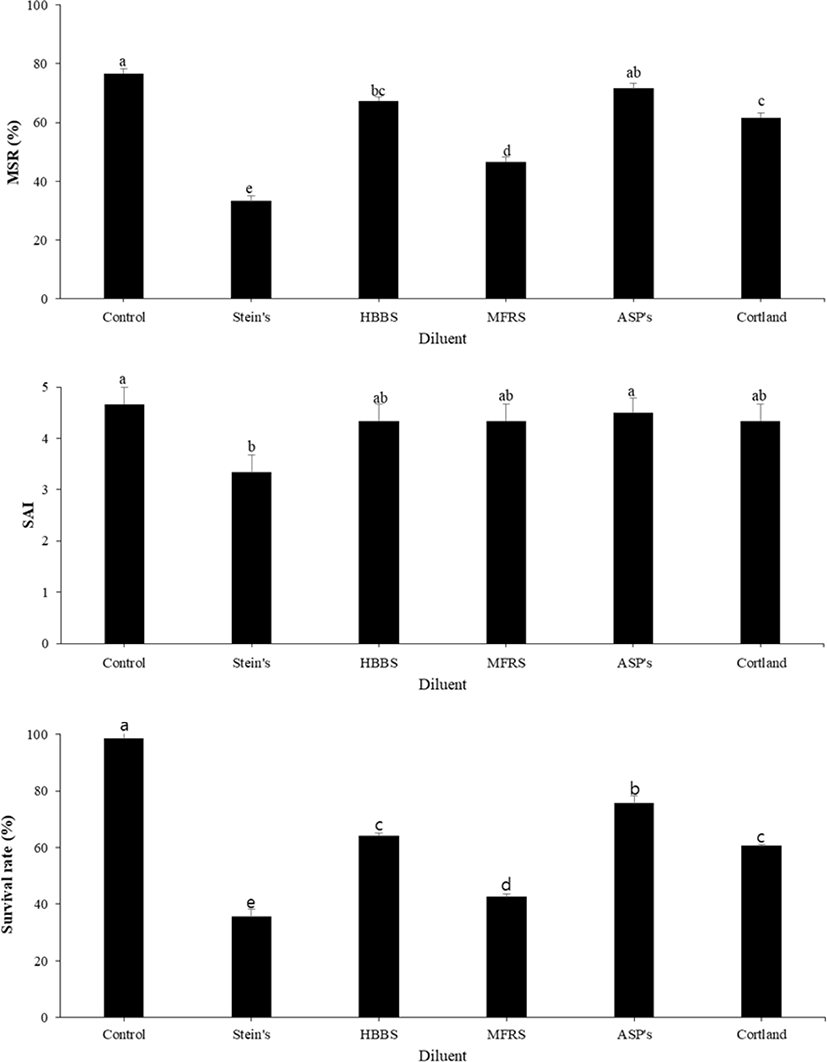
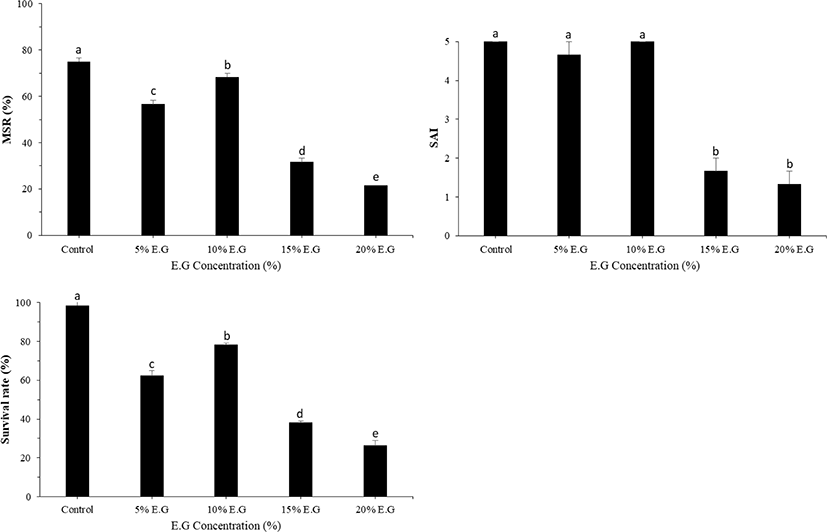
The motility parameters recorded in frozen sperm are provided in Fig. 1. The data obtained indicate a significant effect of the CPA used (p < 0.05) on motility, SAI, and survival rate. Compared with sperm cryopreserved with other CPAs, the best post-thaw motility was recorded for sperm cryopreserved with 10% EG (56.67 ± 2.88%; p < 0.05), followed by that with 10% DMSO (34.33 ± 1.16%; p < 0.05). In contrast, PG, methanol, and glycerol resulted in low MSRs. SAI and survival rates were significantly higher for sperm cryopreserved with EG (4.3 ± 0.58% and 73.33 ± 1.52%, respectively) than for that cryopreserved with other CPAs. When comparing different diluents with 10% EG, ASP of small yellow croaker resulted in the highest MSR (71.67 ± 2.88%), although this value was not significantly different from that obtained using HBBS (67.33 ± 2.51%). SAI and survival rates were significantly higher for ASP (4.5 ± 0.5 and 75.67 ± 2.08%, respectively) than for other diluents (Fig. 2). The effect of various concentrations of EG was investigated using ASP as a diluent. Compared with the use of other concentrations of EG, the highest MSR (68.33 ± 1.66%) and survival rate (78.30 ± 0.42%) were observed using 10% EG (Fig. 3). SAI was not significantly different when 5% and 10% EG were used but was significantly different on comparing 5% and 10% EG and other concentrations.
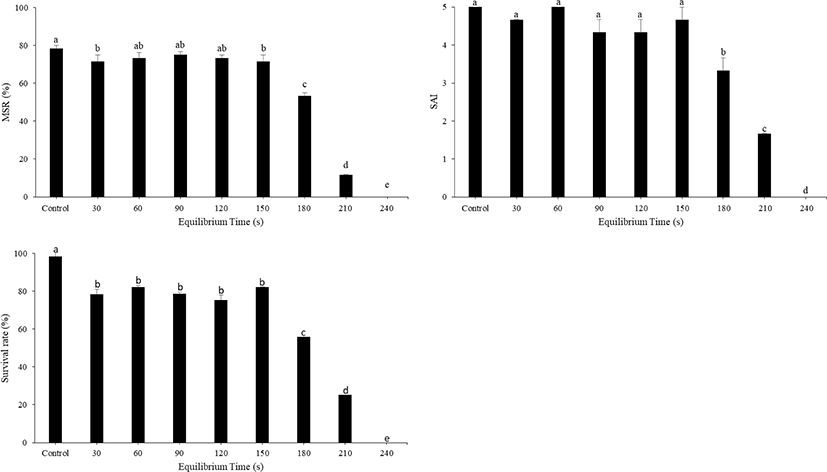
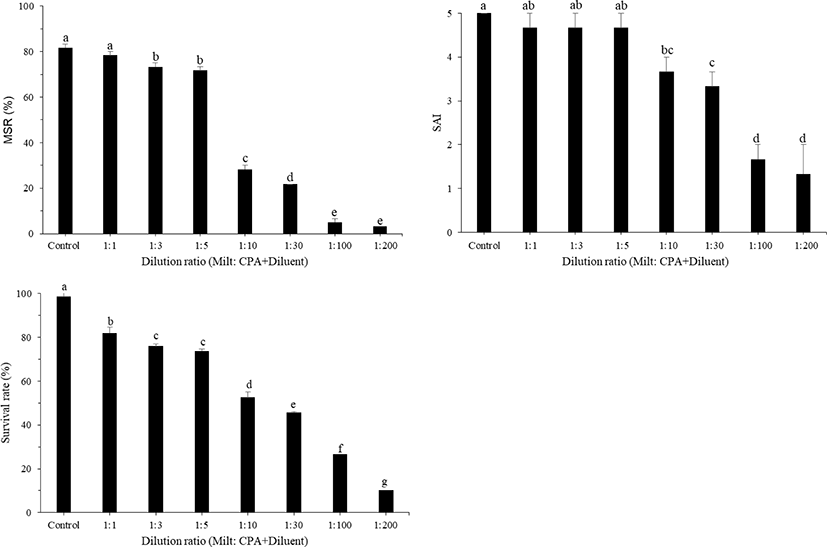
Post-thaw MSR, SAI, and survival rate obtained at various equilibration times during the freezing process are shown in Fig. 4. Post-thaw MSR in large yellow croakers appeared to be stable up to the equilibration time of 150 s, but post-thaw MSR, SAI, and survival rate significantly decreased (p < 0.05) with equilibration times longer than 150 s. The effects of different dilution ratios of sperm to the mixture of CPA + diluent on the motility of post-thaw sperm are shown in Fig. 5. The ratios of 1:1 and 1:3 exhibited the highest MSR at 80.67 ± 0.67% and 79.33 ± 1.20%, respectively, highest SAI of 4.7 for both, and highest survival rates at 95.09 ± 1.02% and 93.97 ± 2.29%, respectively, with no significant difference between both treatments. However, MSR, SAI, and survival rate significantly decreased after treatment with diluent ratios of 1:10–1:200.
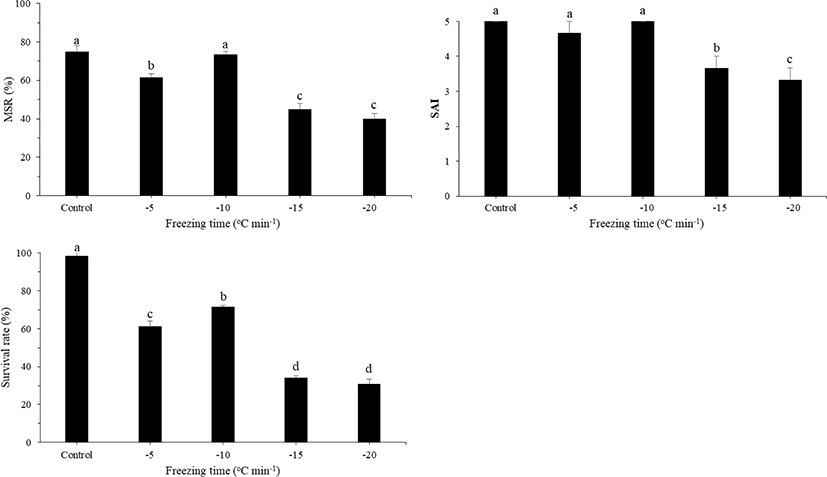
MSR, SAI, and survival rate of post-thaw sperm at different freezing rates during the sperm freezing process are shown in Fig. 6. The highest MSR was observed at the freezing rate of –10°C/min (81.67 ± 1.67%), followed by that at the freezing rate of –5°C/min (79.83 ± 1.04%), with no significant difference between the two treatments. The same freezing rates also resulted in a high survival rate in post-thaw sperm. The SAI value was observed but was no significant difference between the two treatment.
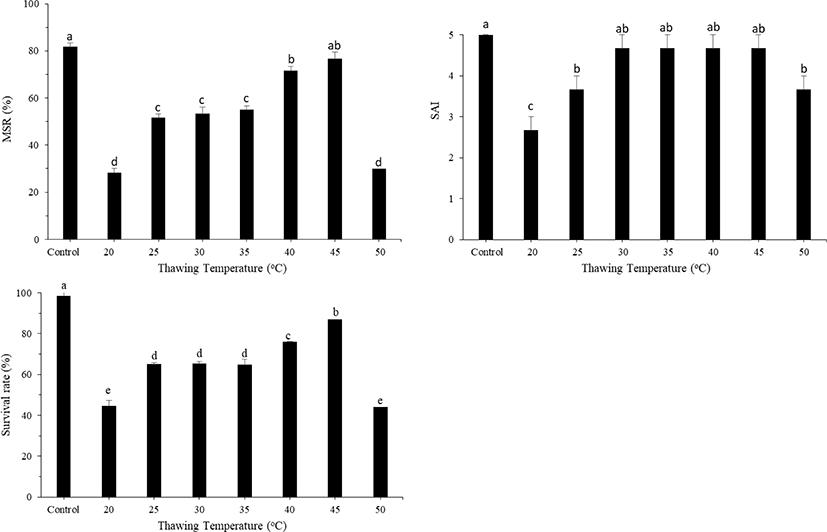
Different thawing methods (20°C, 25°C, 30°C, 35°C, 40°C, 45°C, and 50°C) were applied after sperm freezing in the nitrogen tank (−196°C). The effect of different thawing methods on the post-thaw sperm motility of large yellow croakers are shown in Fig. 7. The best sperm motility was observed after thawing at 45°C for 10 s (76.67 ± 1.67%), followed by that at 40°C (72.33 ± 1.45%), with no significant difference between the two methods. The highest survival rate was observed after thawing at 45°C for 10 s (86.92 ± 0.77%). SAI was significantly lower after thawing at 20°C for 10 s, but there was no significant difference in SAI between treatments at 25°C–50°C.
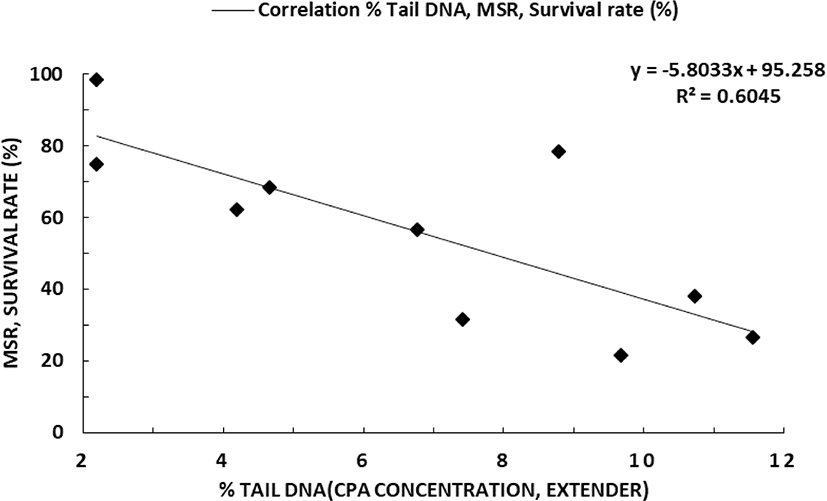
The correlation between large yellow croaker sperm MSR, survival rate (%), and % tail DNA (CPA concentration, Different Diluent) are shown in Fig. 8. The percentage of tail DNA had a negative correlation to MSR and survival rate (y = –5.8033x + 95.258, R2 = 0.6045) with lower % tail DNA damage occurring with higher sperm MSR and survival rate.
To investigate DNA damage following cryopreservation, we observed the effect of different E.G concentrations, with ASP as the diluent, on sperm via SCGE. The highest head length and head intensity values were identified in the control (fresh sperm; 56.04 ± 0.76) followed by 10% E.G (86.71 ± 0.94), which were higher than those obtained with other concentrations of E.G. Further, treatment with 10% ASP showed low levels of tail intensity, percentage of tail DNA, and tail length relative to other treatments but higher levels than those of the control (p < 0.05; Table 3). The effect of different diluents on cryopreservation damages was observed using 10% E.G as the CPA. The highest head intensity and the lowest tail intensity, percent tail DNA, and tail length values were found when ASP was used as the diluent (Table 4).
Discussion
Sperm cryopreservation is one of the most important biotechnological techniques in aquaculture and has many advantages. Understanding the physiology of gametes and the biochemical processes occurring during sperm collection, processing, freezing, and thawing is important for improving sperm cryopreservation techniques (Berlinsky et al., 1997; Cabrita et al., 2005; Suquet et al., 2000). Several factors affect post-thaw sperm motility, including sperm quality, diluent, CPA, equilibration time, freezing rate, dilution ratio, and thawing temperature (Chang et al., 1997). Among these factors, the role of CPA is particularly important. Most cryopreservation injuries can be avoided at high CPA concentrations, but at the same time, the CPA may be toxic to the cells (Bozkurt & Yavas, 2016). In the current study, frozen-thawed sperm motility was evaluated using 10% EG, which is an effective CPA for large yellow croaker sperm cryopreservation. In addition, 10% DMSO exhibited the second best motility ratio in frozen-thawed sperm. This result is similar to that of the study by Jian et al. (2011), who found that there was no significant difference in sperm viability between fresh sperm and frozen-thawed sperm of large yellow croaker when using 5%–20% EG or DMSO as the CPA. According to Xiao et al. (2007), the use of 10% DMSO as CPA showed higher sperm motility in large yellow croakers. In another species of croaker, Le et al. (2011) observed EG to be the best CPA to preserve the quality of frozen-thawed small yellow croaker sperm. In this report, methanol, glycerol, and PG were not suitable in combination with ASP as a diluent because they produced frozen-thawed sperm with low motility. However, some studies have shown glycerol to be an effective CPA in sperm cryopreservation of large yellow croakers (Lee & You, 2002; Xiao et al., 2007). Differences in the effectiveness of CPAs may occur within the same species owing to the differences in the diluents combined with them (Zidni et al., 2020). Moreover, various characteristics of the CPA and the diluent used may affect the maintenance of sperm. Cryoprotectants are important for the protection of the sperm from cold and heat-shock treatments and for the prevention of cell dehydration (Le et al., 2011; Lim et al., 2007). CPAs can also be used to avoid the formation of ice during pre-freezing, but the same levels of CPA can be lethal to unfrozen cells (Cabrita et al., 2010; Chao & Liao, 2001; Cloud & Patton, 2008; Kopeika et al., 2007; Suquet et al., 2000).
Like CPAs, diluents also play an important role in sperm cryopreservation, particularly for long-term cryopreservation. Diluents perform critical functions as regulators of osmotic pressure, pH, and ionic components. In this study, ASP use resulted in high MSR, with no significant difference between ASP-treated cryopreserved sperm and fresh sperm. HBBS and Cortland solution were also good diluents in cryopreservation of large yellow croakers, but SS and MFRS resulted in lower frozen-thawed sperm motility. We found that ASP was the best diluent in this investigation. ASP as a diluent is effective because it is based on yellow croaker seminal plasma (Le et al., 2011; Lim et al., 2007). The use of ASP as a diluent in fish sperm cryopreservation has been extensively investigated for a variety of fish species (Le et al., 2008; Lim & Le, 2013). HBBS and Cortland solution have already been used as diluents for cryopreservation in large yellow croakers and have resulted in good motility of frozen-thawed sperm (Cheng et al., 2015; Jian et al., 2011; Xiao et al., 2007; Xu et al., 2014).
The optimal equilibration time varies among different species and within the same species depending on the type and quantity of CPA/diluent used (Betsy & Kumar, 2020). For effective protection during cooling, sufficient time should be given to facilitate the penetration of CPAs into sperms. In the present study, equilibrium times up to 150 s were effective at maintaining motility at the same level as that of the control. However, decreased sperm motility was correlated with equilibration times longer than 150 s. Sufficient time should be provided for effective protection during cooling to promote the absorption of cryoprotectants into sperms. Because the sperm is sufficiently small and DMSO penetration is rapid, no lengthy equilibration time is required for this CPA (Harvey, 1983).
Dilution ratios could be a significant factor in the efficiency of cryopreservation. In our study, higher motility of the cryopreserved sperm was observed when the ratio of 1:1 was used. Increasing the dilution ratio from 1:10 to 1:200 reduced the percentage of MSR, SAI, and survival rate. A previous study on cryopreservation of large yellow croakers showed higher motility of post-thaw sperm on using the dilution ratio of 1:3 (Jian et al., 2011; Lee & You, 2002; Lin et al., 2006; Xiao et al., 2007). For cryopreservation of fish sperm, the optimum dilution ratio is often represented by a variety of species-specific values from 1:1 to 1:20 (Suquet et al., 2000), and higher dilution ratios generally result in reduced percentage of motile sperm, SAI, and sperm survival rate. The motility duration of black grouper spermatozoa decreased from 40 min to 2 min with an increase in the semen dilution ratio from 1:10 to 1:100 (Gwo, 1993). Increasing the dilution rate from 1:1 to 1:9 did not modify the percentage of motile frozen-thawed turbot spermatozoa (Cosson et al., 1999).
Freezing rate is a significant factor in sperm cryopreservation. In the present study, the optimal freezing rate was in the range of –1°C to –10°C min−1, indicating that these freezing rates could better prevent formation of intracellular ice crystals in sperms during the cryopreservation process than the other rates tested. The best freezing rate for sperm cryopreservation in large yellow croakers was –10°C min−1. Using this freezing rate, intracellular ice crystal formation during cryopreservation was prevented. The optimal post-thaw motility of large yellow croaker was achieved using a freezing rate of 20°C min−1 with HBBS as the diluent and glycerol or DMSO as the CPA (Xiao et al., 2007). Theoretically, to minimize the exposure time of sperm to concentrated extracellular solutions, the freezing rate should be fast enough and yet slow enough to minimize the formation of intracellular ice crystals in sperms to below a damaging level through cellular dehydration. In other marine species, the optimal freezing rate varies among species such as cod (5°C/min), hirame (8°C/min), sea bass (10°C/min), and sea bream (10°C/min) (Fauvel et al., 1998; Mounib, 1978; Suquet et al., 2000; Suquet et al., 2005; Tabata & Mizuta, 1997).
During thawing, the same physiological processes take place in a reverse order as that during freezing. Theoretically, thawing rate should be the same as the corresponding cooling rate. The rate of temperature change must be well moderated to avoid additional stresses due to rates that are either too fast or too slow. In this study, 45°C was an effective thawing temperature, resulting in higher post-thaw sperm motility. According to Xiao et al. (2007), thawing in a 43°C water bath showed optimal post-thaw motility in large yellow croaker. Another experiment, using a thawing temperature ranging from 38°C to 40°C, resulted in higher sperm motility in large yellow croakers than using temperatures outside this range (Lee & You, 2002). In contrast, the optimal post-thaw sperm motility for small yellow croakers was observed when subjected to a 37°C water bath for 30 s (Le et al., 2011; Lim et al., 2010). In other marine species, the optimal thawing temperature is species-specific, with that of the Atlantic halibut, grouper, barramundi, bluefin tuna, and sea bass at 10°C–40°C, 25°C, 30°C, 40°C, and 35°C, respectively (Bolla et al., 1987; Leung, 1987; Withler & Lim, 1982).
In recent years, SCGE has been developed and established as a rapid and accurate system for identifying DNA damage in sperm (Zilli et al., 2003). In the present study, the lowest value of percentage of tail DNA was found in the control treatment (fresh sperm) at 2.19 ± 0.22%, followed by that using 10% EG + ASP treatment (4.66 ± 0.36%), which were significantly different to the values obtained using the other treatments. Investigation of DNA damage in large yellow croaker after cryopreservation using Cortland solution as the diluent and DMSO as the CPA at the concentration of 5%–20% showed no significant differences in motility, comet rate, and damage coefficient between fresh sperm and cryopreserved sperm (Xu et al., 2013; Xu et al., 2014). SCGE results showed that there were no significant differences in DNA fragmentation between fresh sperm and frozen-thawed sperm diluted with 5%–20% DMSO or 5%–20% EG; however, DNA damage to frozen-thawed sperm increased significantly at 25% DMSO or 30% EG (Xu et al., 2014). Significant DNA damage occurred in cryopreserved European sea bass (Dicentrarchus labrax) sperm compared with fresh sperm, but the cryopreservation process did not affect motility or fertilization percentage (Zilli et al., 2003). Similar results were shown by Labbe et al. (2001), wherein DNA stability was lowered by cryopreservation of rainbow trout sperm, but survival rate and quality of sperm remained unaffected. The level of damage to fish sperm DNA is affected by several factors, including the species of fish, its chromosome structure, and the CPA and diluent used for the process of freezing and thawing (Lim et al., 2006; Liu et al., 2014).
Conclusion
In this study, the optimal cryopreservation protocols were determined for large yellow croaker. The use of 10% EG as the CPA and ASP as the diluent resulted in the highest post-thaw sperm motility and the lowest percentage of tail DNA. Post-thaw MSR in large yellow croakers appeared to be stable up to an equilibration time of 150 s and at a dilution ratio of 1:1–1:3. Further, the freezing rate was found to be slow below –10°C/min. Moreover, a thawing temperature of 45°C resulted in the highest post-thaw sperm motility.