
Introduction
Abalone is an economically important shellfish that is extensively farmed worldwide (Gordon & Cook, 2004). The Pacific abalone, Haliotis discus hannai, is the main economically important cultured species in the aquaculture industry in Korea (Park & Kim, 2013); it is regarded as the most commercially important marine mollusk for aquaculture in Korea (KOSIS, 2020). The growth of the abalone aquaculture industry has been accompanied by expansion of the feed industry. Although natural seaweeds, such as Saccharina japonica and Undaria pinnatifida, have been used as abalone feed, there are concerns regarding supply limitations and poor availability in some seasons (Kim et al., 2013). These problems have partially been resolved by the production of commercial abalone feed, generally formulated with S. japonica and U. pinnatifida as the main carbohydrate sources. However, the cost of commercial abalone feed has increased because of unstable S. japonica and U. pinnatifida supply (Hernández et al., 2009; Jang et al., 2018; O’Mahoney et al., 2014). Thus, there is considerable interest in the development of alternative seaweeds and substitute sources to fully resolve these problems.
The brown alga, Sargassum horneri, is a species widely distributed in coastal areas of Korea, Japan, and China (Komatsu et al., 2007; Yoshida, 1963). There is some concern regarding the excessive biomass of S. horneri, which is a major component of floating seaweeds that travel up to hundreds of kilometers over several months on ocean currents (Okuda, 1977; Yoshida, 1963). This excessive biomass represents an emerging environmental problem associated with marine accidents and damage to aquaculture facilities (Smetacek & Zingone, 2013), fisheries, aquaculture organisms (Hwang et al., 2016), and coastal landscapes (Fletcher, 1996; Valiela et al., 1997) in Korea. Although there have been efforts to use S. horneri as a source of bioactive components and agricultural fertilizers or livestock feed, most S. horneri biomass continues to be dumped into landfills or incinerated, causing environmental concerns.
Potential applications of bulk-collected S. horneri could involve industrial processes that utilize all components of the material. There has been some interest in using whole seaweeds, including S. japonica, in the abalone feed industry. Explorations of the potential for using S. horneri as a feed for abalone require assessment of the attractiveness and palatability of S. horneri components for abalone. Abalone can recognize chemical cues transmitted through the water via their chemical detection, tactile, and olfactory organs (Kitagawa & Fusetani, 1989; Uchida et al., 2010). Therefore, the exploration of S. horneri utility as a feed for abalone requires monitoring of abalone behavior and feeding preferences upon exposure to S. horneri components.
Behavioral analysis is necessary for confirmation of an animal’s health and an understanding of the various physiological aspects of that animal’s responses to environmental changes. Aquatic animals are good models for behavioral monitoring because chemical signals can diffuse throughout the homogeneous water environment (Duminda et al., 2021). Numerous factors exist in the aquatic environment; changes can significantly affect an animal’s physiological and behavioral responses (Nagelkerken & Munday, 2016). For some aquatic animals, it is challenging to directly observe and investigate feeding activities because these behaviors may occur under restricted conditions (e.g., during the movement of abalone in the dark). Explorations of the potential for using S. horneri as a substitute source for abalone feed require the development of a system to effectively monitor such behavior, while determining attraction efficiency and feeding preferences. In this study, we identified conditions appropriate for monitoring the movement of Pacific abalone; we also determined that the attractiveness of S. horneri to Pacific abalone was comparable with the attractiveness of standard commercial feed.
Materials and Methods
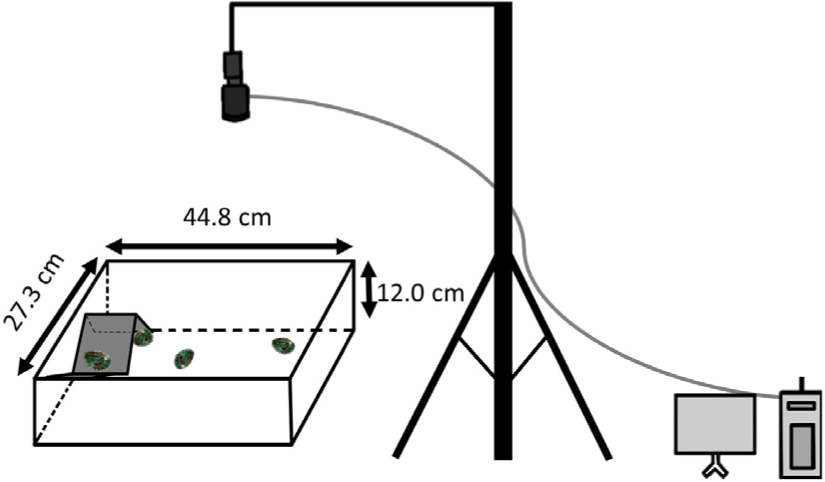
Juvenile Pacific abalone, H. discus hannai, were obtained from SungsilSusan (Wando, Korea). The sizes of H. discus hannai were in the range of shell length 3.2 ± 0.9 cm and body weight 5.9 ± 0.7 g. Water temperature was maintained at 16.8°C–17.5°C with salinity of 31–35 ppt. A rectangular tank measuring 44.8 cm L × 27.3 cm W × 12.0 cm H was separated into an adaptation zone with a shelter and the remaining with test materials. A video camera (Basler ace acA1300-60gm; Basler, Mannheim, Germany) was secured on top of the lighting experiment tank and connected to a computer running tracking software (Ethovision XT 10; Noldus Information Technology, Leesburg, VA, USA) (Fig. 1).
S. japonica and U. pinnatifida were obtained from a local market (Busan, Korea); commercial abalone feed was obtained from Dami Wando Jeonbok Feed (Jeonnam, Korea) (Fig. 2A). Dried seaweed was ground and filtered through a sieve (90-μm mesh). Avicel plates were prepared by pouring 6.25 mL of 20% (w/v) Avicel solution, including 1.77 mL of the material of interest, onto a 90-mm-radius region on glass plates (20 cm × 10 cm). The coated plates were allowed to dry before use.

Eleven juvenile H. discus hannai maintained in a rectangular tank were fed 1% body weight commercial feed daily, but they were starved 1 day before the experiment. To examine the effects of light intensity on the movement of abalone, the following three light intensity conditions were used: fluorescent light at 250 lux (control), dim red light at 15 lux, and dark condition where light intensity was not detectable within device detection limits as 0 lux (Fig. 3). Light intensity was measured using a spectrometer (Sekonic C-7000 SpectroMaster; Sekonic, Tokyo, Japan). Prior to this experiment, the 11 juvenile H. discus hannai were allowed to acclimatize for 5 h under each light condition; subsequently, images were acquired for 6 h. The reliability of the system to monitor movement of H. discus hannai was examined upon the addition of a piece (0.3 g) of S. japonica at the center of the testing region in the tank. Each experiment was repeated three times.

To validate the use of the video tracking system, behavioral analysis was conducted with an Avicel plate containing S. japonica as a positive control and coffee waste as a negative control (Fig. 2B and 2C). The movement of Pacific abalone adapted to dark conditions in the tank was analyzed upon exposure to two Avicel plates containing each material in the detection zone. Abalone movement was recorded over 6 h of exposure to the test materials by a camera secured on top of the tank. Recorded abalone movements were analyzed using Ethovision XT 10 video tracking software (Noldus Information Technology). Heat maps were generated based on the movement patterns of abalone during exposure to each material; the distribution, duration, and frequency of contact with the material zone were calculated, along with latency until first reaction.
Abalone feeding preferences were examined using the powder and methanol extracts of the test materials. Avicel plates prepared as described above were placed in the tank, and the feeding reaction time was examined for 3 h. To compare the palatability of S. horneri with commercial feed, mixtures containing 25%, 50%, or 75% S. horneri (Sh25, Sh50, and Sh75, respectively) were prepared.
Preference analysis was conducted by placing one plate on the corner of the detection zone with another plate on the other side of the tank; the feeding reaction was analyzed for 3 h. The presence or absence of a feed-enhancing effect was expressed as the percentage of Avicel plate area eaten by the abalone.
Results
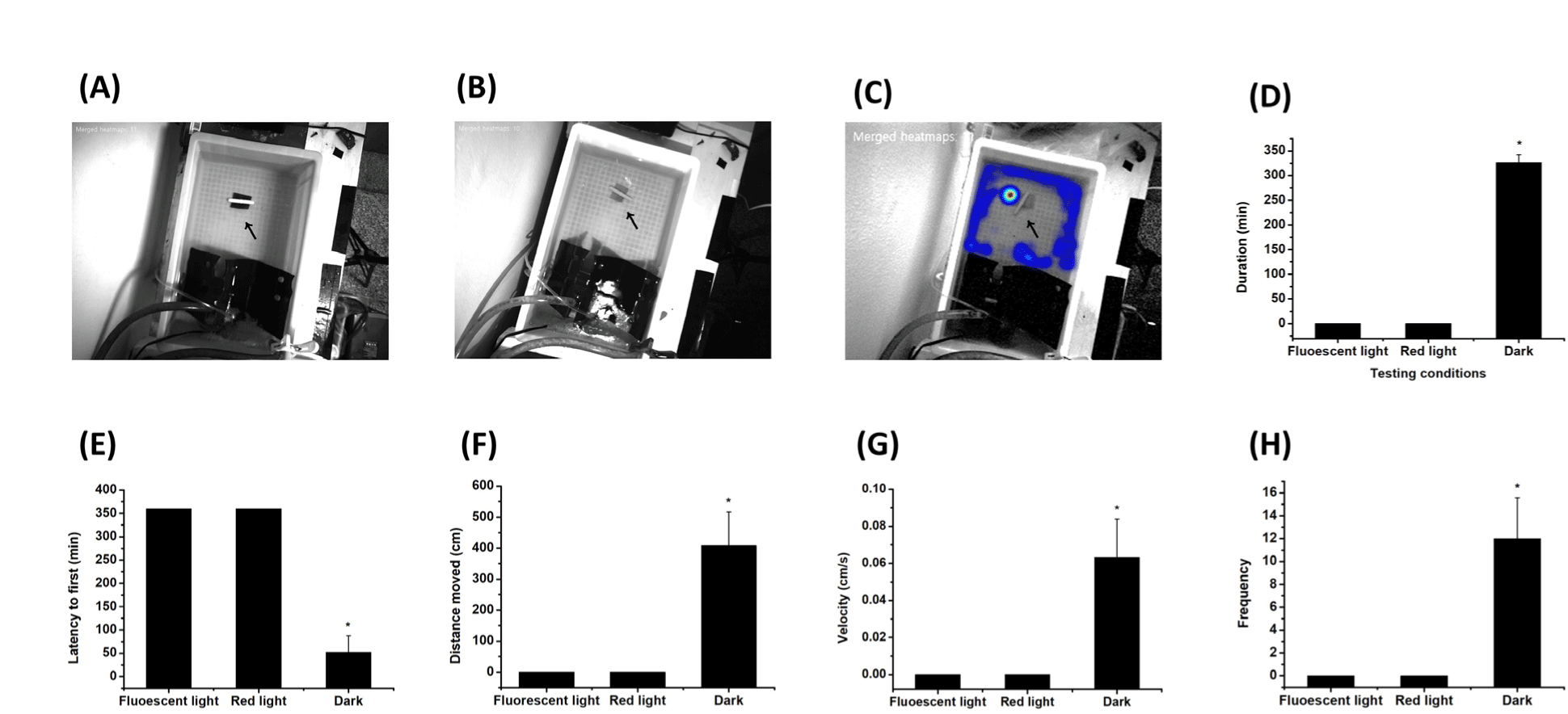
Analysis of animal behavior is important in efforts to identify materials that can attract or repel the organisms. Vision-based monitoring and sonar-based detection systems have been used to detect the movement of marine animals. To establish a camera-based detection system that can monitor the movement of abalone, which mainly move in the dark, there is a need to determine optimal lighting conditions while avoiding excessive exposure to light that would lead to behavioral interference. First, we exposed juvenile H. discus hannai to three different light intensities (250 lux, 15 lux, and 0 lux), then performed movement analysis. Heat map analysis revealed trajectories of Pacific abalone, together with information regarding locations in the tank where they remained for an extended duration. The results of tracking analysis for 6 h indicated no movement of abalone out of the shelter under illumination with fluorescent light or dim red light (Fig. 4A and 4B). However, movement of Pacific abalone around the testing material was clearly detected under the dark (~0 lux) condition, where the wall of the tank was illuminated with dim red light (Fig. 4C). Under these conditions, abalone remained in the zone around the test material for the longest duration, with the shortest latency of 52.2 ± 35.1 min to the first movement. The abalone also exhibited significantly greater movement distance of 407.8 ± 107.9 cm, velocity of 0.06 ± 0.02 cm/s, and frequency of 12.0 ± 3.6 under the dark condition, compared with fluorescent light and dim red light conditions (Fig. 4D–4H). Therefore, the dark condition was sufficient to examine feeding-related behavior of Pacific abalone.
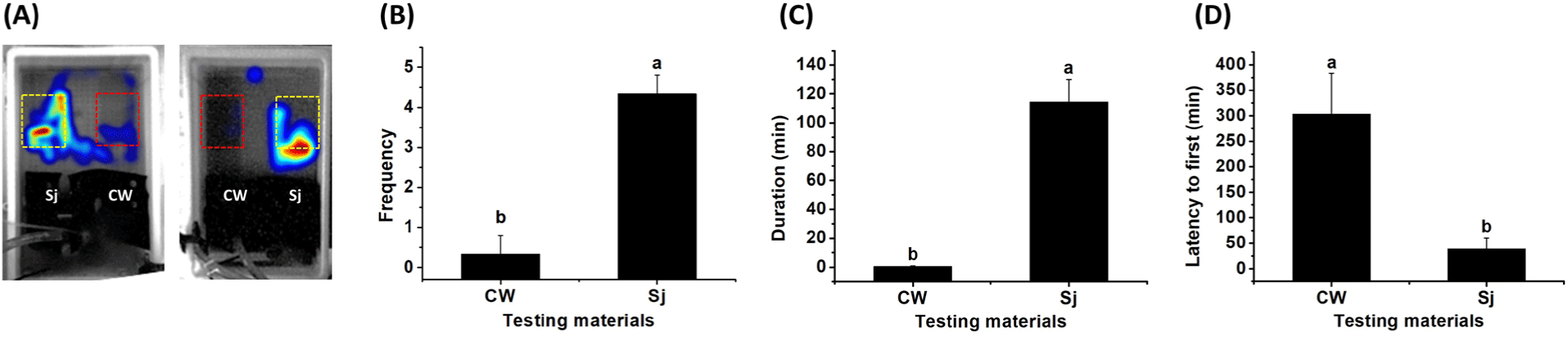
To validate use of the video-based tracking system for analyzing the movement of abalone in response to stimuli, behavioral analysis was conducted using S. japonica, a main component of abalone feed, as a positive control and coffee waste as a negative control; these controls were located at each corner of the detection zone in the tank. Greater movement was detected in the zone containing S. japonica, with considerably less movement in the zone containing coffee waste. To confirm that these results were not related to bias caused by positioning effects in the tank, the experiment was repeated with the materials placed in opposite locations. The results showed a strong attraction effect of S. japonica, but not coffee waste, regardless of position (Fig. 5A). The results also confirmed a significantly higher frequency and duration, and a shorter latency to the first movement of abalone detected in the zone containing S. japonica, compared with the zone containing coffee waste (Fig. 5B and 5C). The latency time was significantly faster for S. japonica than for coffee waste (Fig. 5D). These results confirmed that our system is appropriate for behavioral analysis; it does not exhibit any biasing effect concerning the location of the zone in the tank.
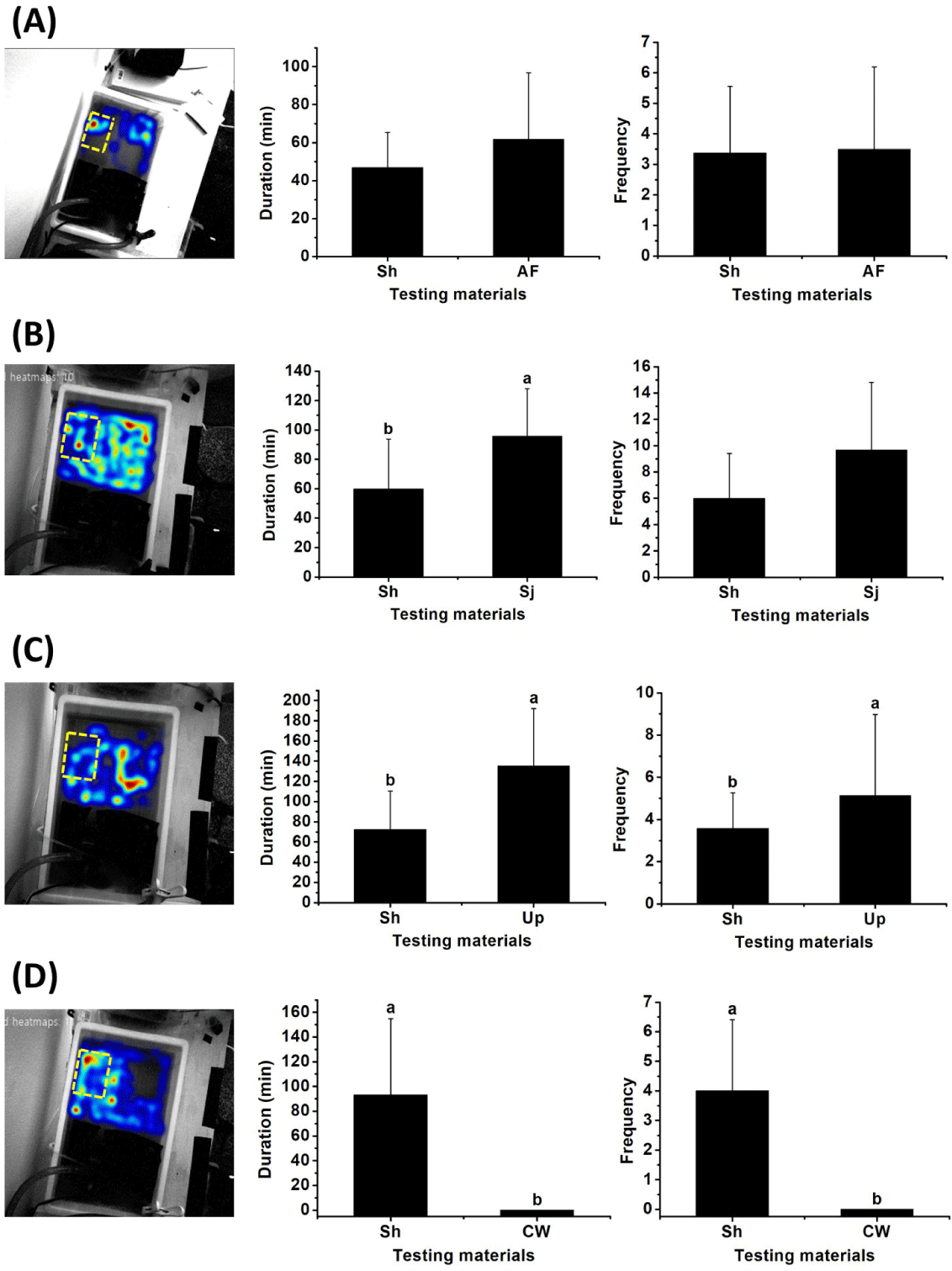
There has been concern regarding S. horneri, which causes extensive damage to aquaculture facilities and fisheries. Explorations of the potential for using S. horneri as feed for abalone in aquaculture require assessment of its attractiveness in comparison with other feeds. To compare the degree of attraction toward S. horneri, movement of Pacific abalone was examined upon exposure to an Avicel plate containing S. horneri along with exposure to another plate containing S. japonica, U. pinnatifida, commercial abalone feed, or coffee waste. The levels of attraction were first investigated by heat map analysis, tracing the patterns of movement toward test materials (Fig. 6). The results were quantitatively analyzed according to the frequency and duration of attraction of H. discus hannai to each material. Heat map analysis indicated that S. horneri had similar attractiveness to commercial abalone feed; there were no differences in the frequency (3.4 ± 2.2) or duration (47.0 ± 18.5 min) of Pacific abalone presence near S. horneri compared with its presence near commercial feed (3.5 ± 2.7 and 61.8 ± 35.0 min, respectively, both p > 0.05) (Fig. 6A). Although there was no significant difference in the frequency of Pacific abalone presence around S. horneri and S. japonica (6.0 ± 3.4 vs. 9.7 ± 5.1, p > 0.05), the duration was significantly shorter for S. horneri than for S. japonica (59.8 ± 34.0 vs. 95.7 ± 32.5 min, p < 0.05), implying that S. japonica had greater attractiveness (Fig. 6B). U. pinnatifida was also significantly more attractive than S. horneri (frequency 5.1 ± 3.8 vs. 3.6 ± 1.7, p < 0.05; duration 135.3 ± 56.7 vs. 72.3 ± 37.9 min, p < 0.05) (Fig. 6C). The reliability of the results was confirmed by comparison with coffee waste as a negative control (Fig. 6D).
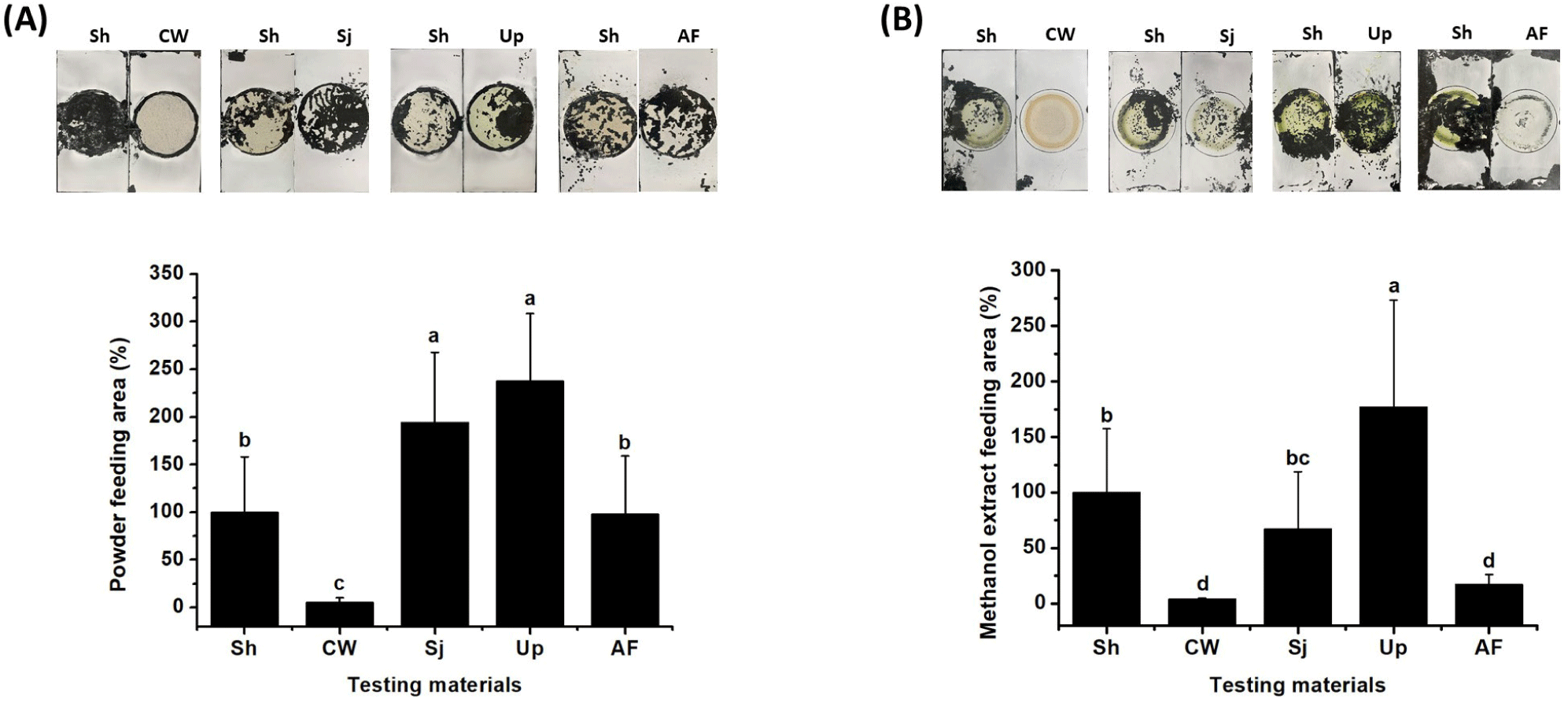
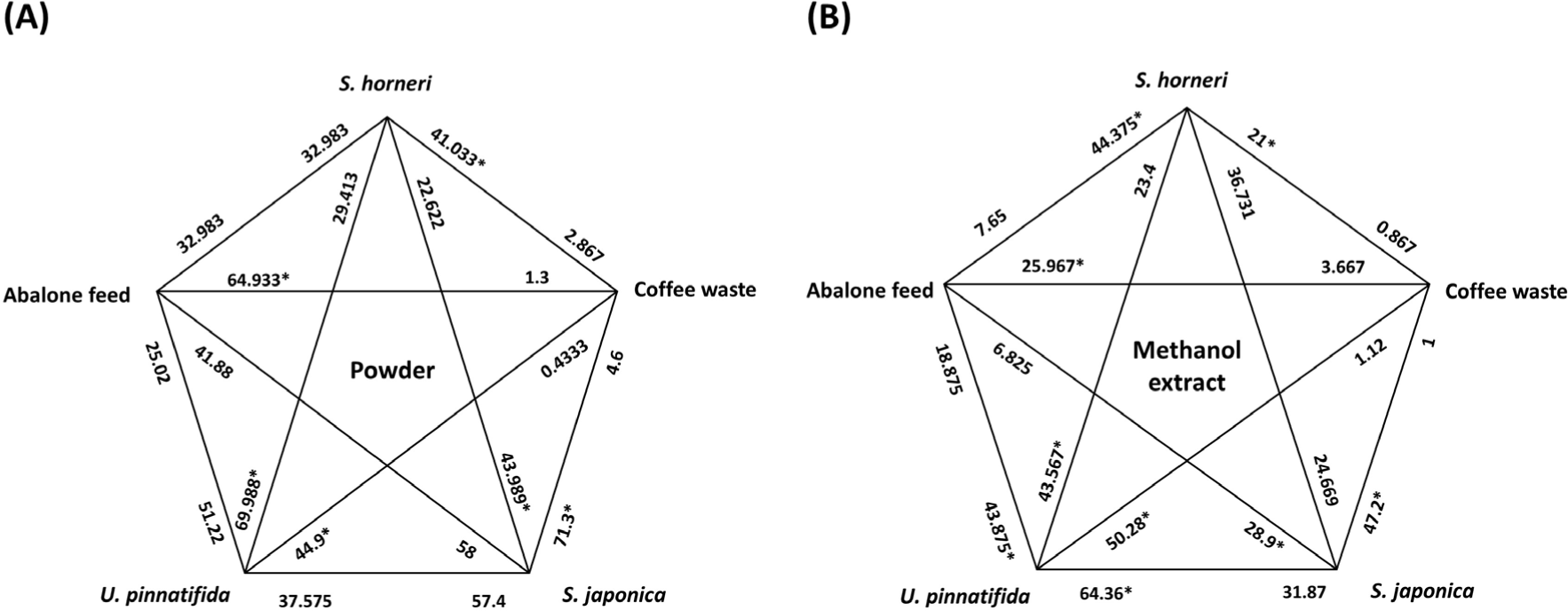
To validate the feasibility of using S. horneri as feed, attraction efficacy and actual feeding preference must be evaluated. Experiments were conducted with one Avicel plate containing S. horneri powder and another Avicel plate containing powders of S. japonica, U. pinnatifida, commercial abalone feed, or coffee waste in the tank. Feeding preference was determined by analyzing the area of the Avicel plate eaten by Pacific abalone, relative to the area of the S. horneri plate that was eaten (regarded as 100%). The highest preference levels were observed for U. pinnatifida (238.0 ± 70.6%) and S. japonica (194.4 ± 73.3%) followed by S. horneri (100 ± 49.0%) and commercial feed (99.7 ± 63.6%); there was no preference toward coffee waste as a negative control (7.0 ± 4.6%) (Fig. 7A). Thus, the preference of abalone for S. horneri was similar to the preference for commercial feed. Preference analysis was also conducted with methanol extracts of the test materials to determine the chemical and physical characteristics of the components that attracted Pacific abalone. For this analysis, the methanol extract of each test material was applied to the designated area of each Avicel plate, and the plates were used after they had been allowed to dry. Although abalone exhibited a significantly greater preference for the methanol extract of U. pinnatifida (177.3 ± 96.2%) than for the methanol extracts of S. horneri (100 ± 57.4%) and S. japonica (67.2 ± 51.7%), its preference for the methanol extract of commercial feed was similar to its preference for the methanol extract of coffee waste (17.2 ± 8.9% and 4.1 ± 0.6%, respectively, p > 0.05) (Fig. 7B). The overall preferences of abalone for each material and their methanol extracts were compared based on these experimental results (Fig. 8). The results showed that the preference for S. horneri was comparable with the preferences for other materials that have been used as feed for Pacific abalone in aquaculture.
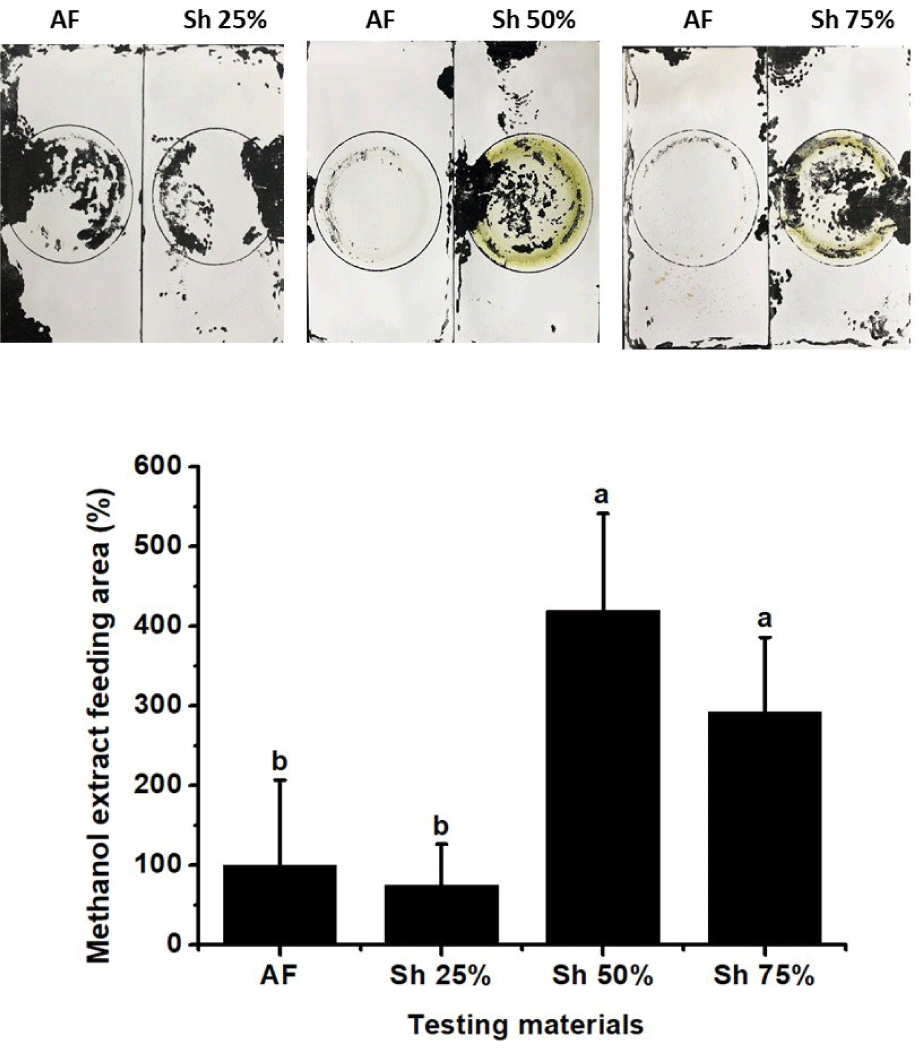
To further examine whether components of the methanol extract of S. horneri can be used to enhance the palatability of commercial feed, experiments were conducted with mixtures of commercial feed and 25%, 50%, or 75% (w/w) S. horneri extract (Fig. 9). Although there were no significant differences in preference for powdered formulas or the addition of 25% methanol extract of S. horneri, significant increases in preference were observed with addition of 50% and 75% S. horneri methanol extract. These results indicated that components present in the methanol extract of S. horneri could be used as a feed additive to enhance feed preference for Pacific abalone.
Discussion
Movement, an integral component of animal activity, is essential for foraging, attraction, aggression, and antipredation behaviors. Thus, there is a need to monitor the ability to move, which can strongly influence survival (Almbro & Kullberg, 2008; Domenici, 2001; Weber, 2009). Whereas it is easy to conduct vision-based monitoring of diurnal animals, it is challenging to analyze the movement of nocturnal animals (e.g., abalone), which mainly move and feed in the dark. Haliotis fulgens reportedly exhibited the highest growth rate under conditions of constant darkness; the survival rate and feed intake of Haliotis corrugata larvae were much higher in the dark than under conditions of constant light (Gorrostieta-Hurtado et al., 2009). In the present study, we first investigated appropriate lighting conditions for use in monitoring the movement of Pacific abalone, an important aquaculture species in Korea. We examined whether the movement of Pacific abalone could be monitored using a camera under fluorescent light (250 lux), dim red light (15 lux), and dark conditions (~0 lux) with indirect lighting from dim red light. H. discus hannai tends to prefer dark environments and longer wavelengths of red and orange light (Gao et al., 2016); as expected, no distinguishable movements (e.g., feeding activity) were observed under fluorescent light or dim red light conditions. However, dark conditions with indirect dim red light appeared to be sufficient for camera-based visual detection of Pacific abalone movement without affecting their nocturnal behavior. The reliability of the system was confirmed using S. japonica included in feed for Pacific abalone along with Laminaria spp. and Undaria spp. (Bangoura et al., 2014) as a positive control, and coffee waste as a negative control. To confirm that the observed preference toward S. japonica but not coffee waste was not related to a zone bias effect, the experiment was conducted with the same materials in the opposite positions in the tank. The results confirmed a clear preference toward S. japonica regardless of position; it was also confirmed that our video tracking system under indirect dim light illumination could be used to monitor abalone movement and evaluate abalone attraction to various materials. Therefore, this method was used to compare the attraction efficacies of seaweed materials, including S. horneri, for Pacific abalone. The assay was focused on S. horneri, which is associated with environmental problems in aquaculture facilities (Fletcher, 1996; Ha et al., 2016; Smetacek & Zingone, 2013), because there is a need to find effective uses for this material. Notably, the composition of seaweed changes according to season and place of growth, and the composition differs among parts of the thallus (Ishii et al., 1980; Nishide et al., 1988); thus, it is challenging to determine feeding preferences based on the results of experiments using seaweed collected at a single site. However, S. horneri is presumed to possess an attractant for Pacific abalone. Our results showed that the attractiveness of S. horneri was comparable with the attractiveness of commercial feed, suggesting that S. horneri can be used as a feed additive.
We also explored the potential attraction efficacy of S. horneri in comparison with other materials, including S. japonica, U. pinnatifida, and commercial feed, to validate its efficacy as a feed additive for enhancing palatability to Pacific abalone. The preference was confirmed based on the area on the Avicel plate eaten by Pacific abalone (Kawamura et al., 2001). Feeding tests with powdered materials revealed the highest preferences for U. pinnatifida and S. japonica, followed by similar levels of preference for S. horneri and commercial feed; there was no preference for coffee waste as a negative control. Although S. horneri has been identified as a source of vitamins, protein, fiber, polyunsaturated fatty acids, and both macro- and microbioactive compounds (Ortiz et al., 2006), Sargassum spp. have been utilized only in fertilizers and animal feeds (Kang, 1968). Furthermore, the growth rate of juvenile abalone was reportedly higher upon addition of a mixture of U. pinnatifida and S. horneri in comparison with either single treatment alone (Ansary et al., 2019). These observations, together with our findings regarding feeding preference toward S. horneri, suggest that it can be used as an abalone feed substitute.
To examine the chemical and physical characteristics of components of S. horneri, preference tests involving methanol extracts were also performed. The results showed the following order of feeding preference: U. pinnatifida > S. horneri > S. japonica > commercial feed > coffee waste. The results indicated a greater preference for the methanol extract of S. horneri than for the methanol extracts of S. japonica and commercial feed, despite similar levels of preference toward their powdered forms. These differences may have been related to the lower content of fat-soluble substances in the powdered forms. However, studies on Haliotis discus reeve demonstrated high levels of feeding stimulation by fat-soluble extracts of S. japonica, sea lettuce, and U. pinnatifida (Lee & Kim, 2013) in which a mixture of chloroform and methanol (C:M = 2:1) in contrast to 100% methanol used for extraction in the present study. It was also reported that complex lipids such as digalactosyl diacylglycerol and phosphatidyl choline among the fat-soluble substances extracted from wakame (Undaria sp.) stimulate feeding of H. discus reeve juveniles (Kitagawa & Fusetani, 1989). This was consistent with our results indicating that the methanol extract of S. horneri contains substances that attract H. discus hannai and stimulate its feeding. The results suggest a way to efficiently use large quantities of S. horneri biomass causing environmental problems in aquaculture facilities.
To further explore whether components in the extract of S. horneri could be used to enhance the palatability of commercial feed, we mixed commercial feed with increasing amounts of S. horneri extract up to 75% (w/w). The results showed a minimal increase in feeding preference upon the addition of 25% S. horneri extract, but significant increases in palatability were observed with the addition of 50% and 75% S. horneri extract. These results suggested that the methanol extract of S. horneri contains components with potential uses in the enhancement of commercial feed palatability.
In summary, a video tracking system with a suitable lighting condition was established to detect the feeding-associated movement of Pacific abalone together with Avicel-based feeding preference analysis. Feeding preference analysis of Pacific abalone for various seaweeds (e.g., S. horneri, Saccharina japonica, and Undaria pinnatifida) and commercial abalone feed indicated that the attractiveness of S. horneri was comparable with the attractiveness of commercial feed, suggesting that the biomass of S. horneri can be used as a feed additive. Our result also showed that the methanol extract of S. horneri contains components with potential uses in the enhancement of commercial feed palatability.