
Introduction
Freshwater bodies are invaluable ecosystem maintaining com-plex habitats. A remarkable variety of biodiversity, from microscopic to large organisms, is supported by these dynamic habitats. Streams and rivers are freshwater environments that are essential for sustaining a variety of aquatic organisms and ecological balance (Allan & Flecker, 1993). These waterbodies are valued for their exceptional biological richness and for offering a variety of ecosystem services. Stream fish are an important part of the diverse range of organisms that live in these dynamic environments. They play a significant role in both the overall biota including fish, particularly those found in tropical rivers, have still not been well studied in terms of their habitat preferences, functional characteristics and distribution patterns. Tropical rivers represent biodiversity hotspots, harboring a plethora of fish species with unique habitat requirements. These rivers support a variety of ecosystems rich in life because of their warm temperatures, high levels of sunshine, and plentiful rainfall (Ochieng et al., 2019). Many fish species live in their waters, and they differ greatly in size, shape, behavior, and ecological roles.
The habitat preferences of streams are influenced by various factors, such as water depth, water velocity, cover, and substratum composition (Pringle, 2003). Moreover, stream fish have a variety of adaptations that show how well they can live in particular ecological niches (Poff & Zimmerman, 2010). Studies have shown that fish preferences are largely determined by factors like substrate composition, water velocity, and hydrological conditions (Kennard et al., 2007; Ugbor et al., 2023). These habitats are delicately shaped by the interaction of several factors, including temperature, nutrition availability, substrate composition, and water flow. This results in a complex assembly of fish species preferences and adaptations (Poff et al., 1997). Furthermore, changes in hydrological conditions affect the heterogeneity of fish habitats and alter fish species diversity. Fish habitats and their ability to navigate rivers can be impacted by changes in water flow parameters, such as depth and velocity. In a river, shallow areas can make it difficult for fish to pass, especially for larger species, while deeper sections can offer protection and appropriate habitat (Guo et al., 2018).
The Upper Awash River in Ethiopia, characterized by a network of major tributaries, stands as a pivotal aquatic ecosystem, hosting a diverse array of stream fish. However, the river is exposed to different human activities, including waste released from domestic and factory sources, land degradation, water resource depletion, rapid population growth, and urbanization (Lakew & Moog, 2015). Garra is one of the small indigenous fish species in the Awash River. Understanding the community structure and habitat preferences of this fish is essential for effective conservation and sustainable management of this vital freshwater resource. In Ethiopia, in terms of community structure, more emphasis has been given to commercially exploited fish species, with little effort made for small indigenous species like the genus Garra (Englmaier et al., 2020; Erarto, 2020; Tesfay et al., 2019; Wubie et al., 2017). In addition, information on habitat selectivity in relation to changes in environmental variables is limited. Therefore, the primary objective of this study is to investigate the diversity and abundance of fish at the mesohabitat level in the upper Awash River while concurrently exploring their intricate interrelationships with pertinent environmental variables.
The study of fish distribution at the habitat level may assist environmental managers in developing policies for river resource management at the local and regional levels. In addition, knowing the community dynamics of stream fish has broader implications for the adaptability and health of ecosystems (Pringle, 2003). It is also an essential step in understanding the distribution patterns of fish. Studying fish habitat preferences allows us to anticipate how these changes might affect fish distributions and populations.
Materials and Methods
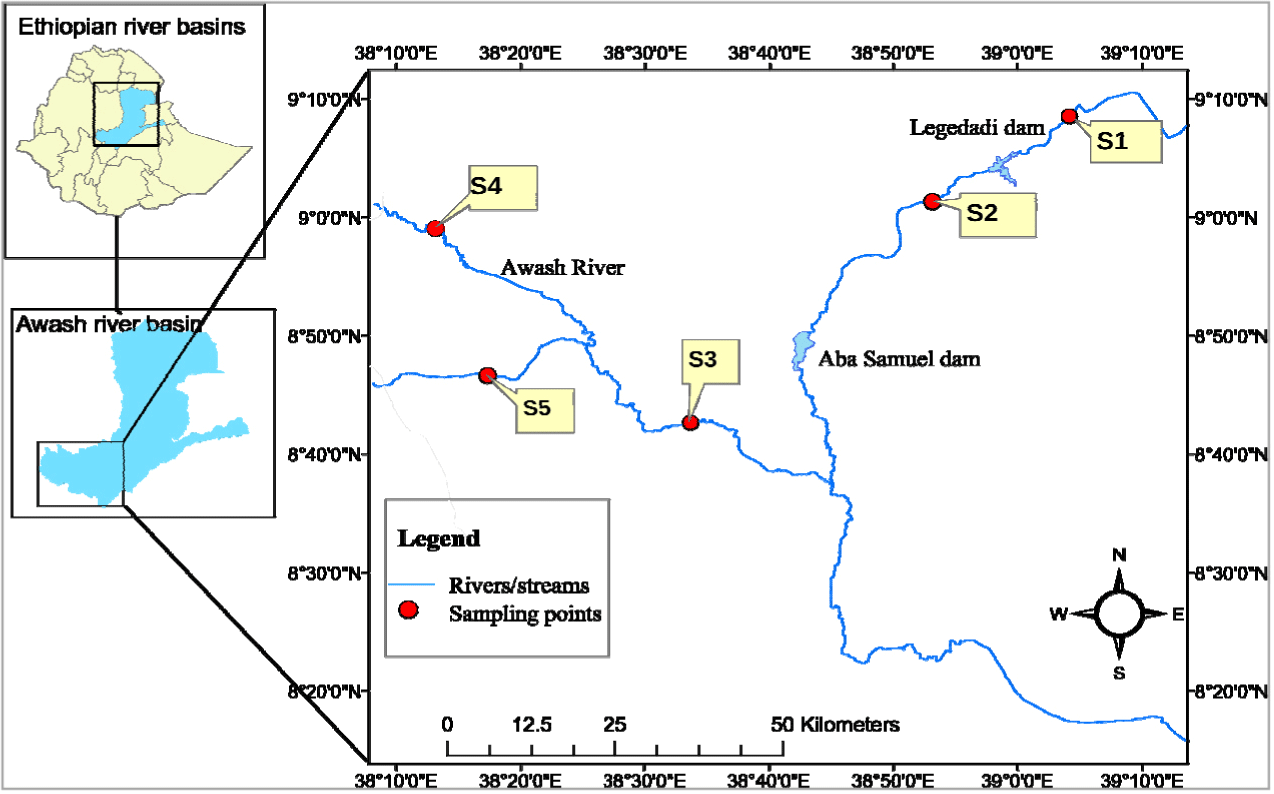
The study was conducted between May and October 2021 in the upper Awash River, covered distinct seasonal phases: May (pre-rainy dry season), June to August (rainy season), and September to October (post-rainy dry season). The Awash River basin, originating from the Ethiopian central highlands, is known for being one of the largest river basins. It flows northeast through the Rift Valley, eventually reaching the Ethiopian-Djibouti border, where it meets the salty Lake Abbe, situated at an altitude of approximately 250 m. The length of the Awash River spans approximately 1,200 km, encompassing a total coverage of 112,700 km², and serves as a home for around 10.5 million individuals. The upper region of the Awash River covers an area of 10,748 km², with tributaries originating from various directions in Showa that flow to the point where it flows into Lake Koka. A total of five sampling sites were selected, including two sites from the main river Awash and three sites from its tributary streams, namely Legedadi and Teji (Fig. 1). The selection of these study sites was carefully planned in alignment with the study objectives, ease of sampling access, fish presence, and the specific habitat types, namely pool and riffle. To ensure comprehensive data collection, sampling was carried out in both a pool and a riffle habitat at each of the sampling sites. Pool habitats were characterized by deeper water with slower flow, while riffle habitats were characterized by shallower water with faster flow over substrate features. Legedadi above the dam (S1) is the first site with low human activity and better canopy cover. The second site (S2) is a dam effluent receiving site situated downstream of the Legedadi Dam. The Awash Kunture (S3) site is the third site with high water flow and depth, whereas the fourth site (S4) is near Ginchi town, where intensive farming activities are expected to be a source of pollution. The last site, Teji (S5), is a hydrologically altered site due to water diversion for irrigation purposes.
The temperature (°C), dissolved oxygen (mg/L) and conductivity (µS/cm) of water were measured in situ using a multi-probe system (YSI 556MPS, YSI, Yellow Springs, OH, USA). Water depth was measured using tape mounted on a stick and an echo-sounder (DIR-60A, Lenco, Kerkrade, Nederlands) was used at the pool of the River Awash Kunture (S3), which was the deepest site in the study area. The floatation method was used to measure stream velocity by measuring the time it takes a floating material (orange) to travel a known distance. Each of the parameters was taken three times at each sampling site and the average value was considered in the final analysis. A two-liter of water sample was taken from each sampling site, stored in ice-cooled boxes and transported to the limnology laboratory of the National Fishery and Aquatic Life Research Center (NFALRC), Sebeta, Ethiopia. The nutrient concentration was determined following the standard method (APHA, 1998). Habitat conditions in each mesohabitat type at each sampling site were assessed using a visual scoring system following O’Brien et al. (2011). We classified the dominant substrate types, namely bedrock, boulders, cobbles, gravel, sand, mud, and silt. Habitat parameters also included dominant cover types that comprised root wads, vegetation and undercut banks (Kleynhans, 1999).
Fishing was done from each habitat type (pool and riffle) using an electrofisher (Model EFGI 1300 with a single anode array and pulsed DC 60 Hz, voltage 1–470 V) in a zigzag pattern against the water flow. The net barrier was set at the upstream and downstream ends to prevent fish from escaping. Fishing was conducted in a zigzag pattern against the water flow in both the riffle and pool habitats (Gozlan et al., 1999). To minimize harm caused by the electric field, fish caught were promptly transferred to buckets filled with river water and stored until identification and counting were completed. On-site fish identification was conducted following Damesé (2012). Unidentified fish were collected in a plastic container with 10% formalin and transported to the NFALRC for confirmation and validation. Any fish identified in the field were returned to their respective habitats. A two-pass removal method was employed following Seber & le Cren (1967).
Habitat preferences of the species were determined by evaluating species distributions across different habitat types, specifically pool and riffle habitats, as well as sites with varying environmental conditions (e.g., high water volume vs. effluent-receiving sites). Species abundance and diversity were analyzed in relation to key environmental variables such as water depth, velocity, substrate composition and environmental variables.
The analysis of variance (ANOVA) was used to compare differences in environmental variables and fish community structure between different habitat types using the function aov in R software (R Core Team, 2018). Only if it was significant, a Tukey’s HSD test be performed to get the differences between any two different habitats using the function Tukey HSD in R. Principal component analysis (PCA) was used to observe the association between fish and environmental variables in different habitats. Fish diversity and evenness were calculated based on the equations given (Equations (1) and (2)).
where pi = (ni/N) * (ni/N), ni = number of specimens per taxon, N= total number of individuals per site.
where, H’ max represent the maximum possible diversity of the site and ln (S) for the natural logarithm of species.
Results
From the environmental parameters measured, total phosphorous, nitrate, depth and velocity showed significant variations among sampling sites (ANOVA, p <0.05; Table 1). The highest water velocity and depth were recorded at the S3 site, while the lowest values were recorded at the hydrologically altered site, S5. The site with agricultural activity, S4, had higher nutrient concentrations (TP, total phosphorus and NO3), whereas the dam effluent receiving site, S2, had a higher conductivity value.
The substrate and benthic cover compositions of each habitat are presented in Table 2. Riffle habitat was dominated by gravel, cobble, and boulder substrates while, pool habitat was dominated by silt, sand, and mud substrates. There were nearly equal amounts of benthic cover in both habitats.
During the present study, we recorded a total of 1,784 fish, representing seven species belonging to the family Cyprinidae, across all sites. The riffle habitat had a higher number of fish compared to the pool habitat. Specifically, sites S1 and S3, which were characterized by relatively higher water depth and flow velocity, had a higher number of fish in their riffle habitat, whereas the pool habitat had fewer fish. On the other hand, the remaining sites with shallow water had a higher number of fish in their pool habitat compared to the riffle habitat (Table 3).
In terms of fish species distribution, both mesohabitats in all sampling sites of the upper River Awash were highly dominated by small indigenous Garra species. Other commercially exploited fish species, namely Oreochromis niloticus, Cyprinus carpio, and Clarias gariepinus, were less abundant and restricted only to the pool habitat type (Table 3).
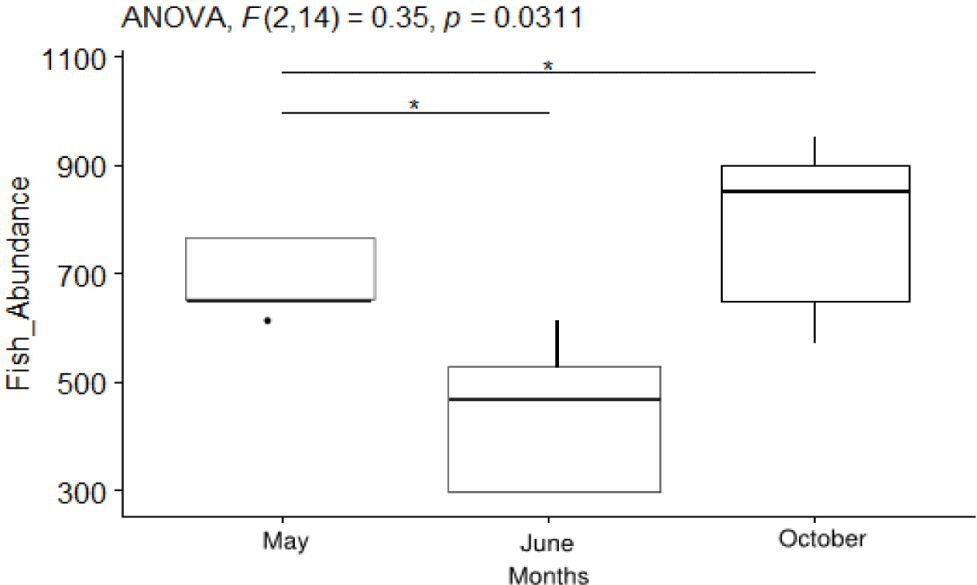
The monthly variation in fish abundance is presented in Fig. 2, showing average ± SE values. ANOVA revealed significant variation in fish abundance across sampling months (F [2, 14] = 0.35, p =0.037). The highest fish abundance was observed in October, while the lowest was recorded in the rainy month of June. Post-hoc tests indicated significant differences between May and June (F [2, 14] = 0.35, p =0.019) and between June and October (F [2, 14] = 0.35, p =0.022; Fig. 2).
Based on the present study, the Shannon diversity index ranged from 0.52 to 1.45, with higher average diversity reported in pool habitat of S3 site, with high water volume. Riffles had lower diversity, with an average value of H’ = 0.87 (Table 4). The species evenness varied from 0.44 to 0.92, where the maximum value was obtained in the pool of S2, dam waste receiving site.
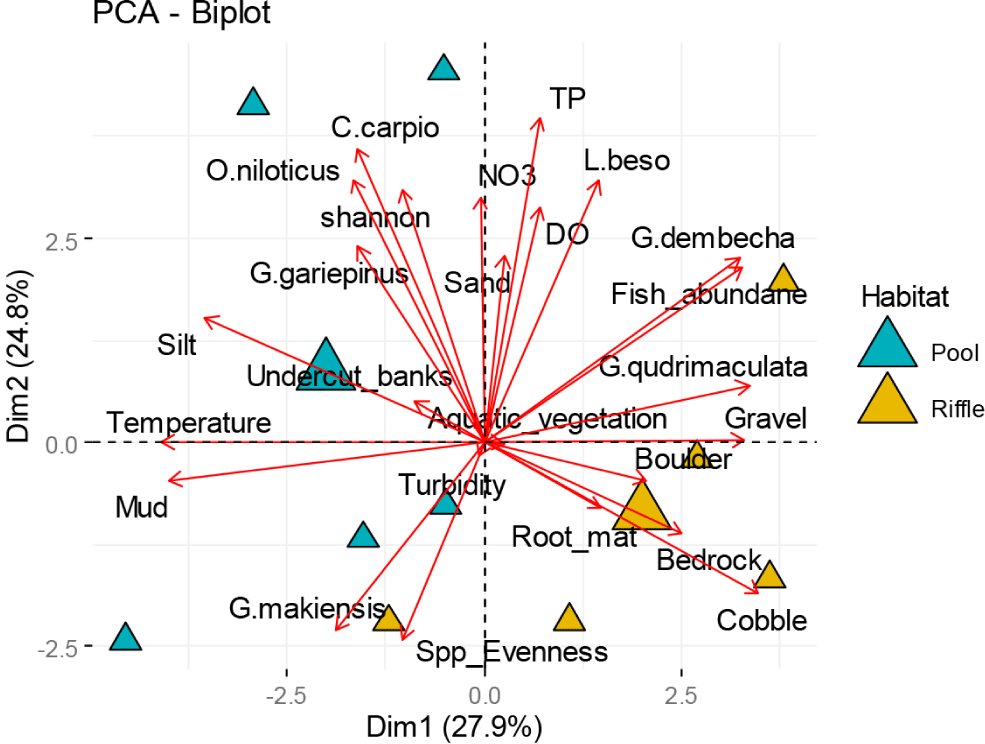
Fig. 3 shows the relationship between habitat variables and fish community structure. Indigenous fish species, such as Garra dembecha, Garra quadrimaculata, Garra makiensis, and L. beso, showed a positive correlation with riffle habitat. This habitat was characterized by dominant environmental variables like TP, nitrate, temperature, gravel, boulders, root mats, bedrock, and cobble. On the other hand, commercially important fish species (O. niloticus, C. carpio, and C. gariepinus) were closely associated with pool habitat, which was characterized by undercut banks, silt, mud and sand. Shannon diversity showed a positive relationship with pool habitat, while species evenness and fish abundance were positively correlated with riffle habitat.
During the study period, fish were divided into two size categories: small (2.5 to 12.5 cm) and large (12.6 to 22.6 cm; Fig. 4). The majority of fish caught (1,280) fell into the small size class accounting for 71.75% of the total, while the large size class contributed 504 fish (28.25%). Analyzing habitat types, it was observed that small fish were more commonly found in pool habitats with slow-moving water, although this difference was not statistically significant (t-test, t600 = 0.13, p >0.05). Conversely, large fish were more abundant in riffle habitats (t-test, t250 = 0.021, p <0.05).

Discussion
The observed differences in environmental characteristics between the sampling sites offer important information on how human activity affects water quality. Previous studies have confirmed that significant anthropogenic modifications have disrupted natural flow dynamics (Guo et al., 2018), as evidenced by the lowest depth and water velocity recorded at site S5 due to river diversion for irrigation farming. The higher levels of TP and nitrate (NO3–) at the site with agricultural activity (S4) are consistent with previous studies highlighting the substantial impact of agricultural runoff on nutrient enrichment in water bodies (Xia et al., 2020). Intensive farming methods, such as fertilizer application and land disturbance increase nutrient loads and accelerate the eutrophication processes in aquatic environments (Carpenter et al., 1998; Gotkowska-Płachta et al., 2016). The observed higher conductivity value at the dam effluent receiving site, S2, is consistent with expectations, reflecting the influence of anthropogenic inputs such as waste discharge from the wastewater treatment plant and urban runoff, which typically increase conductivity due to dissolved ion content.
During the present study, a total of 1,784 species were collected, dominated by cyprinids, like most rivers experienced in Ethiopia. G. dembecha and G. quadrimaculata were dominant in all sampling sites exposed to different anthropogenic stressors (Table 3), indicating the tolerance of Garra species to different levels of pollution as reported in the previous studies (Englmaier et al., 2020; Wubie et al., 2017). L. beso was found in S1, slightly impacted sites, and S5, shallow and hydrologically altered sites. However, it was totally absent at S2, a highly impacted dam effluent receiving site. This is in line with Englmaier et al. (2020), who reported the absence of L. beso at polluted sites with high conductivity values. G. makiensis was totally absent at the high velocity and water volume sites. Similarly, Englmaier et al. (2020) noted, G. makiensis showed preference for intermediate water velocity and avoided high water flow velocity.
The results presented in this study provide valuable insights into the fish population dynamics and habitat preferences within the upper River Awash. The comparison between riffle and pool habitats reveals interesting patterns with riffle habitats generally harboring a higher number of fish, particularly Garra species that favor fast-flowing water conditions (Getahun & Stiassny, 1998). Furthermore, the dominance of Garra species in all sampling sites indicates the species’ ecological importance in the upper River Awash. The Garra species’ preference for riffle habitat may be explained by their well-known adaptation to lotic environments, where they frequently live on rocky substrates with fast moving water (Getahun, 2007). The preference of Garra species for riffle habitats was further supported by the positive correlation between higher abundance and water velocity, as documented by Erarto et al. (2020) in the same region. In general, the higher number of fish in riffle habitat aligns with previous findings on lotic ecosystems (Poff et al., 1997). Higher dissolved oxygen concentrations, faster flow rates and more substrate variability are characteristics of riffle environments that support a wide variety of fish species (Statzner & Bêche, 2010).
Interestingly, commercially exploited species such as Oreochromis niloticus, Cyprinus carpio, and Clarias gariepinus were predominantly associated with pool habitats. This result is in line with studies suggesting that pool habitats, which are defined by deeper water and slower flow, frequently offer larger fish species providing a refuge and possibilities for feeding (Harvey & Stewart, 1991).
Water depth and velocity play a crucial role in determining fish assemblages. It was found that fish had a tendency to aggregate in larger numbers in riffle habitats, especially in sites (S1 and S3) that have deep and fast water flow. This might be attributed to the presence of boulders and rock substrates in the riffle, which act as resistance to slowing down the river water’s flow. As a result, this resistance keeps food from being carried away by powerful currents. Previous research has shown similar trends, with habitats including a higher percentage of boulder and rock substrates showing higher fish abundances (Mohd-Azham & Singh, 2019). In contrast, the depth of riffle habitats in sites S4 and S5 was found to be shallow, rendering them less favorable for fish, attributed to increased exposure to predation. Additionally, streams are dynamic environments that can undergo alterations in flow regimes, including drying up and forming isolated pools during the dry season (Taylor & Warren, 2001). Specifically, the Tji stream (S5) represents a small stream that has been modified to divert water for irrigation purposes, leading to changes in water flow. As a result, fish prefer pool habitats with higher water volumes over riffles. This also might be attributed to the comparatively higher water temperature found in shallow stream riffles, which causes fish to migrate to pool reaches that provide cooler water and serve as thermal refuges (Tate et al., 2006). Our study found that substrate composition and benthic cover influenced the distribution of fish in pool and riffle habitats. Riffle habitats supported a greater number of fish than pool habitats. This distinction can be attributed to differences in substrate and cover composition between these two mesohabitat types. Our PCA ordination analysis revealed a positive relationship between the majority of fish community variables and riffle habitat, particularly in areas with coarser substrates such as cobble, boulder, and rock. This finding supports previous research (Hilling et al., 2021; Taylor et al., 2019), which found that substrate types such as cobble had a significant influence on fish abundance in rivers. In addition, riffles were found to support higher densities of benthic macroinvertebrates than pools. This can be attributed to the presence of diverse substrate types in riffles, which make them important areas for fish food production (Gordon et al., 2004). In our study, the dominant fish species were from the genus Garra, which are bottom feeders that graze on gravel or rock substrates. The availability of such substrates in riffle habitats is likely to have contributed to this species’ aggregation in those areas. Furthermore, habitat heterogeneity was found to influence fish distribution. For example, the S2 and S5 sites, characterized by limited vegetation cover and less habitat complexity, registered lower numbers of fish than other sites (Table 3).
The Shannon diversity index was relatively higher in the pool habitat than the riffle. One possible explanation for this phenomenon is that fish species prefer to spend more time in deep pools, which require less energy for swimming and foraging than navigating fast-moving currents. Our finding is consistent with previous studies (Erarto, 2020; Tesfay et al., 2019; Wubie et al., 2017). Other studies have demonstrated that water depth has a significant impact on fish distribution and community composition (Guo et al., 2018). However, species evenness was found to be lower in pools than in riffles, implying that the pool habitat is preferred by the majority of taxonomically similar species.
The relationship between water volume, habitat conditions, and species diversity is well-documented in ecological studies. In the present study, site with greater water volume, S3 supports higher species diversity, likely due to more favorable habitat conditions. Increased water volume enhances habitat heterogeneity, providing diverse niches that support a wider range of fish species. For example, studies have shown that fish species richness is positively correlated with hydrological attributes such as water depth and dissolved oxygen levels (Kumar et al., 2025). Conversely, sites that receive effluent discharge often experience reduced fish diversity due to negative impacts on water quality. The introduction of pollutants can lead to altered nutrient levels and oxygen depletion, creating stressful conditions for many fish species. Research indicates that effluent discharge significantly affects fish assemblages, favoring more tolerant species while diminishing sensitive ones (Hoyer and Gehebe, 2014).
The observed relationship between fish size and the nature of the two habitats, riffle and pool, can be attributed to the interaction of habitat characteristics with the physiological and ecological traits of fish species. Small fish, ranging in size from 2.5 to 12.5 cm, are the most abundant in pool habitat. This might be attributed to the fact that the fine sediments in pool habitat provide ideal spawning grounds and shelter for small-bodied fish, which are better suited to maneuvering in slower-flowing, more stable environments. Lower water velocity in pool habitats reduces energy expenditure for smaller fish, allowing them to thrive and exploit available resources efficiently (Polivka, 2020). Additionally, the accumulation of organic matter in pool habitats can support higher levels of primary productivity, sustaining populations of small prey organisms that serve as food sources for smaller fish species (Huang et al., 2019). This finding is supported by the research conducted by Singh & Agarwal (2013) in the Mandakini River, India, and Erarto (2020) in the Gumara River, Ethiopia, which confirmed the preference of small-sized fish for pool habitats with slower water velocity. Large-sized fish, on the other hand, prefer riffle habitat capable of withstanding high-water velocity. Cobble, rock, and gravel substrates dominated the Riffle habitats (Table 3), providing ample hiding places and refuge from predators, allowing larger fish to establish territories, and reducing predation risk. Furthermore, riffle habitats frequently have higher water velocities and oxygen levels, providing ideal conditions for larger, more active fish species that can navigate and forage in these environments (Statzner & Bêche, 2010). The abundance of benthic invertebrates associated with the complex substrate increases food availability for larger fish, promoting growth and survival.
The observed temporal fluctuations in fish abundance highlight the complex relationship between environmental variables and fish population dynamics in riverine ecosystems. Our findings reveal a notable decline in fish abundance during the peak of the rainy season, particularly in June, which is consistent with existing literature highlighting the disruptive impacts of heavy rainfall on fish populations (Morgan et al., 2004). Heavy rainfall can cause floods and increase flow velocities, destroying habitat and displacing fish populations. The increase in water turbidity and rapid flow velocities can impair fish feeding efficiency and heighten their vulnerability to predation (Winemiller & Jepsen, 1998). The resulting habitat disruption and diminished foraging opportunities ultimately contribute to reduced fish abundance in affected areas. Furthermore, intense rainfall events may cause downstream displacement of fish populations, exacerbating the decline in abundance observed in the study area (Morgan et al., 2004). This downstream displacement phenomenon emphasizes the interconnectedness of riverine ecosystems and the potential for localized disturbances to have a cascading effect on fish populations across the watershed.