
Introduction
The increasing use of sunscreens and cosmetic products for ultra violet (UV) protection has raised considerable concerns regarding the environmental accumulation and contamination of organic UV filters, which are key ingredients in these products (Molins-Delgado et al., 2016;Tao et al., 2023). Owing to their high lipophilicity, many UV filters are resistant to degradation, persist in aquatic environments, and have the potential to bioaccumulate in aquatic organisms (Fent et al., 2010; Gago-Ferrero et al., 2012).
Among various UV filters, 2-ethylhexyl 4-methoxycinnamate (EHMC) is one of the most commonly used chemical UV absorbers worldwide (Cadena-Aizaga et al., 2020; da Silva et al., 2022). Regulatory agencies have imposed concentration limits on EHMC in cosmetic products (FDA, 2021; MFDS, 2024). Recently, the Scientific Committee on Consumer Safety (SCCS) raised concerns regarding the potential endocrine-disrupting properties of EHMC in various organisms (SCCS, 2025), and the U.S. classified EHMC as a category III ingredient, indicating insufficient data to confirm its safety and effects (FDA, 2021).
Due to its low water solubility and widespread use, EHMC has been frequently detected in aquatic environments worldwide. Reported concentrations include up to 4,043 ng/L in coastal waters of Hong Kong, 1,080 ng/L at Japanese beaches, and 390 ng/L in Norwegian coastal areas (Langford & Thomas, 2008; Sankoda et al., 2015; Tsui et al., 2014). Accumulating evidence indicates that EHMC exerts adverse effects on fish, particularly during early developmental stages. Previous studies have demonstrated that EHMC induces developmental abnormalities and hatching delays in zebrafish embryos (Yang et al., 2024), while parental exposure increased malformation and mortality rates in F1 embryos (Zhou et al., 2019a). Furthermore, EHMC exposure altered the gene expression of neurotoxicity-related proteins, such as myelin basic protein and growth-associated protein 43 and reduced glutathione levels, a key antioxidant involved in cellular protection (Chu et al., 2021; Yang et al., 2024).
Endocrine-disrupting effects of EHMC have also been reported in adult fish. Exposure of female fathead minnows (Pimephales promelas) to EHMC resulted in suppressed final oocyte maturation and significant downregulation in gene expression of 3β-hydroxysteroid dehydrogenase (3β-HSD), a key enzyme involved in steroid hormone synthesis (Christen et al., 2011). In adult zebrafish, EHMC exposure significantly decreased estradiol-17β and vitellogenin levels while increasing testosterone levels, indicating anti-estrogenic and androgenic activities (Zhou et al., 2019b).
Although monitoring the ecological effects of UV filters ideally requires full life-cycle assessments of marine fish, such studies are limited due to the difficulties associated with inducing natural spawning and maintaining marine species under laboratory conditions. Consequently, most toxicity assessments of EHMC have relied on freshwater model species, such as zebrafish and medaka (Lin et al., 2016; Wang & Cao, 2021), while studies focusing on marine fish remain scarce. Given the distinct physicochemical differences between freshwater and seawater, including pH, salinity, and hardness, the toxicity of chemical contaminants may differ substantially between these environments (Leung et al., 2001). Therefore, the development of suitable marine fish models is essential for accurately assessing the ecological risks of EHMC in marine ecosystems.
The chameleon goby (Tridentiger trigonocephalus), a coastal fish species well suited for laboratory experiment, was selected as the experimental model in this study. Our previous studies have established laboratory conditions for natural spawning of this species and reported reproductive cycle, early gonadal development, and sex differentiation (Cho et al., 2014; Hwang & Baek, 2013). The objective of this study was to investigate the toxic effects of EHMC on the embryonic development of the chameleon goby (T. trigonocephalus), a coastal marine fish species. Specifically, we aimed to: (1) characterize morphological abnormalities induced by EHMC exposure during embryogenesis; (2) evaluate sublethal endpoints including yolk sac utilization, heartbeat rate, survival, and hatching success; and (3) assess the suitability of the chameleon goby as a marine model species for ecotoxicological research on UV filters.
Materials and Methods
Adult chameleon gobies (11 females and 10 males) were collected using hand nets from Namcheon and Minrak Harbor, Suyeong-gu, Busan, Korea, in May 2025. Mean body length and weight of collected fish were 5.7 ± 0.6 cm and 4.3 ± 0.4 g, respectively. Captured fish were immediately transported to the laboratory and acclimated in glass aquaria (75 × 45 × 45 cm) with artificial seawater. Acclimation and maintenance followed the protocol described by Hwang (2011). In brief, after a 2-week acclimatization period, spawning was induced by adjusting the photoperiod in 2-week increments to 10L:14D for weeks 0 to 2, 12L:12D for weeks 2 to 4, and 14L:10D for weeks 4 to 6, with a water temperature of 20.0 ± 0.5°C. To induce spawning, broodstock were maintained under controlled conditions at a water temperature of 20.0 ± 0.5°C, salinity of 30 ± 1 psu, and a photoperiod of 14L:10D.
Fish were fed a commercial pellet diet (Tetra Bits®, Melle, Germany) to apparent satiation twice daily. Polyvinyl chloride (PVC) pipes (5 cm in diameter and 15 cm in length) were placed in the tanks as spawning substrates. Tanks were inspected daily for observation of spawning. Six days later, fertilized eggs attached to the inner surfaces of the PVC pipes were observed and carefully collected using fine forceps with minimal physical damage. Approximately 2,000 fertilized eggs were collected within 6 hours post-fertilization for use in EHMC exposure experiments.
EHMC was purchased from Sigma-Aldrich (CAS no. 5466-77-3, purity ≥ 98%, St. Louis, MO, USA). Dimethyl sulfoxide (DMSO; CAS no. 67-68-5, Sigma-Aldrich) was used as the solvent. DMSO at 0.1% (v/v) has been widely used as a carrier solvent in fish embryo toxicity studies without significant adverse effects on development, survival, or hatching success (Fong et al., 2016). Fertilized eggs within 6 h post-fertilization were randomly distributed into 6-well plates, with 30 embryos placed in each well containing filtered artificial seawater. Exposure concentrations of EHMC were set at 5, 50, and 500 µg/L, and the final concentration of DMSO was maintained at 0.1% (v/v) in all treatments, including the control with only DMSO addition. Preliminary tests confirmed that this DMSO concentration did not affect survival, hatching success, or morphological development (data not shown).
Embryos were exposed under the same incubation conditions described above, and the experiment was terminated at 144 h post exposure (HPE), when hatching in the control group was complete. To maintain nominal exposure concentrations, 80% of the rearing water was changed every 24 h. Each treatment was conducted in triplicate.
To evaluate the effects of EHMC on embryonic development, ten embryos per well were randomly selected for measurements of yolk sac volume, heartbeat rate, occurrence of malformation, survival, hatching success and notochord length of newly hatched larvae on schedule (Table 1).
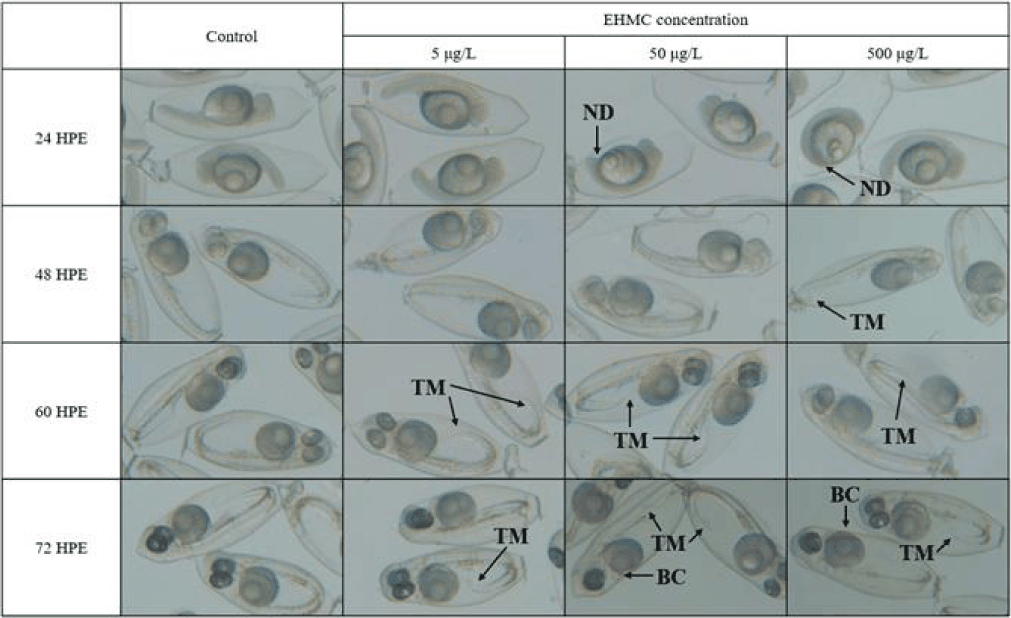
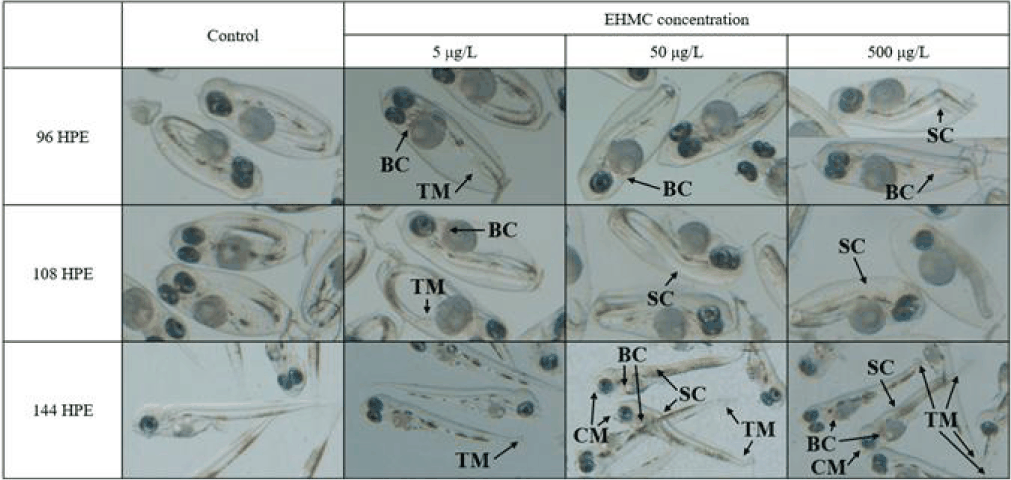
Morphological observations were conducted at 12 h intervals using a stereomicroscope equipped with a digital imaging system (Discovery V12, ZEISS, Oberkochen, Germany). The terminology of malformations is as follows: tail malformation (TM): any deviation from the normal linear extension of the tail region, including bending, curling, or incomplete separation from the yolk sac; blood congestion (BC): abnormal accumulation of blood cells (visible as dark reddish pooling) in the pericardial region or along the trunk vasculature, in contrast to free-circulating blood in control embryos; non-detachment of the tail (ND) was scored separately at 24 HPE as the failure of the tail to physically separate from the yolk mass; spinal curvature (SC): visible lateral or dorsoventrally bending of the vertebral axis compared to the straight-axis reference in control embryos; craniofacial malformation (CM): any visible deformity of the head region, including misshapen or reduced jaw, abnormal eye position, or asymmetric craniofacial structure, compared to control embryos at the same developmental stage.
Yolk sac volume was measured at 24 h intervals after the fusion of multiple oil droplets into a single yolk mass. Yolk sac volume was calculated using the following equation: Yolk volume (mm3) = π / 6 × yolk length (YL) × yolk height (YH)2. Heartbeat rate was measured after completion of cardiac formation, at 72 HPE. Heartbeats were counted for one minute and expressed as beats per minute (bpm). Survival rate was calculated as the percentage for the number of live embryos to the initial number of fertilized eggs (30 per well). Hatching success was calculated as the percentage of hatched larvae from survived embryos. Notochord length of newly hatched larvae was measured to the nearest 0.01 mm.
All data are presented as mean ± SEM from three replicates (n = 3) in each measurement. Statistical analysis was performed using IBM SPSS Statistics (version 29.0, IBM, Armonk, NY, USA). Normality was assessed using the Shapiro–Wilk test. For normally distributed data, one-way analysis of variance (ANOVA) was conducted followed by Tukey’s post hoc test. For data that did not meet normality assumptions, the Kruskal–Wallis test was applied, followed by the Mann–Whitney U test to compare treatment groups with the control. Statistical significance was set at p < 0.05.
Results
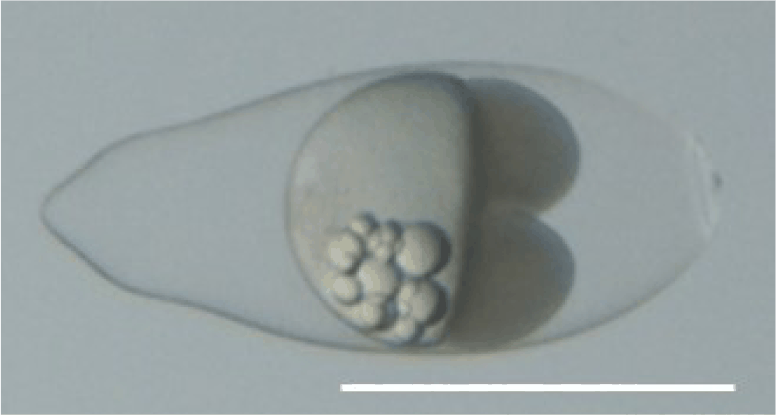
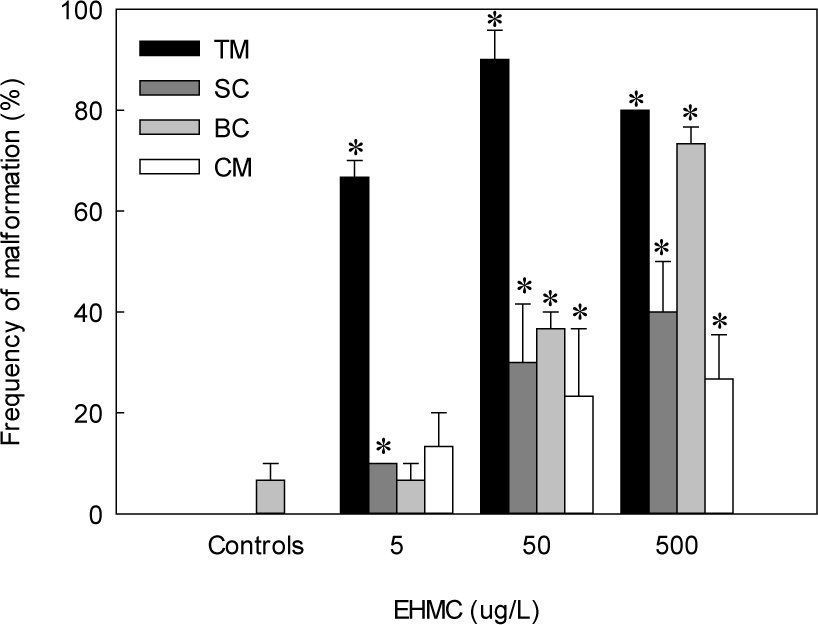
The fertilized egg of chameleon goby was ellipsoidal in shape, transparent, demersal, adhesive and contained multiple oil droplets (Fig. 1). The embryos exposed to 50 and 500 µg/L exhibited non-detachment of tail from the yolk sac at 24 HPE (Fig. 2). TM was first observed at 500 µg/L of EHMC at 48 HPE, and was subsequently observed in all treatment groups by 60 HPE. At 72 HPE, the embryos exposed to 50 and 500 µg/L demonstrated BC in the pericardial region. At 96 HPE, BC was observed in all treatment groups, and SC was observed at 500 µg/L of EHMC (Fig. 3). At 108 HPE, SC was also observed in the embryos exposed to 50 µg/L of EHMC. At 144 HPE, CM was observed in the embryos exposed to 50 and 500 µg/L of EHMC. During the exposure period, TM was the most frequently observed abnormality, and the incidence of BC increased with increasing EHMC concentration (Fig. 4).
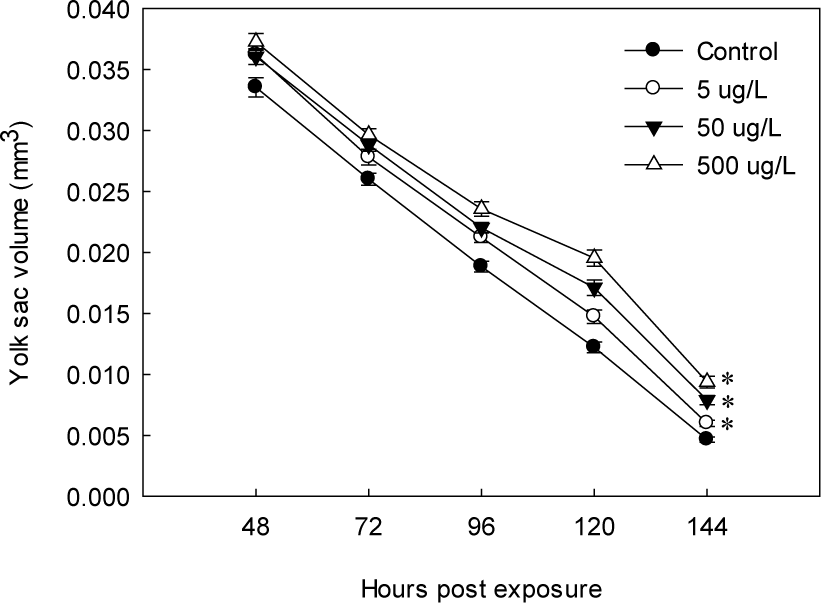
Yolk sac volume decreased in all treatments as time elapsed; however, EHMC exposure significantly delayed yolk absorption (Fig. 5). At 144 HPE, yolk sac volumes in all treatments were significantly larger than those of the control, indicating impaired yolk utilization. The inhibition of yolk absorption exhibited a clear concentration-dependent pattern.
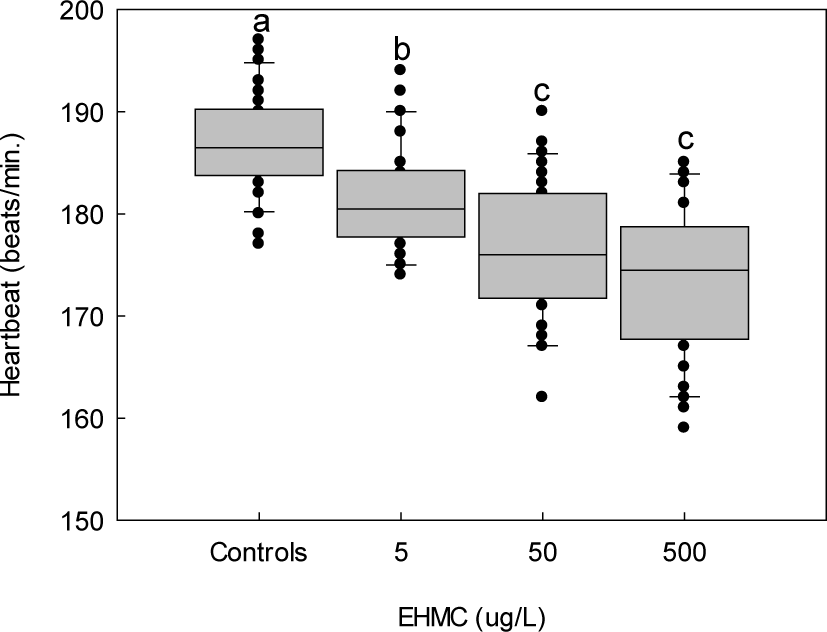
Heartbeat rates measured at 72 HPE decreased significantly with increasing EHMC concentration (p < 0.05). Heartbeat decreased as the concentrations of EHMC increased. Mean heartbeat rates were 186.8 ± 4.9 bpm in the control, compared to 181.6 ± 5.4, 176.6 ± 6.7, and 173.3 ± 7.5 bpm at 5, 50, and 500 µg/L, respectively (Fig. 6).
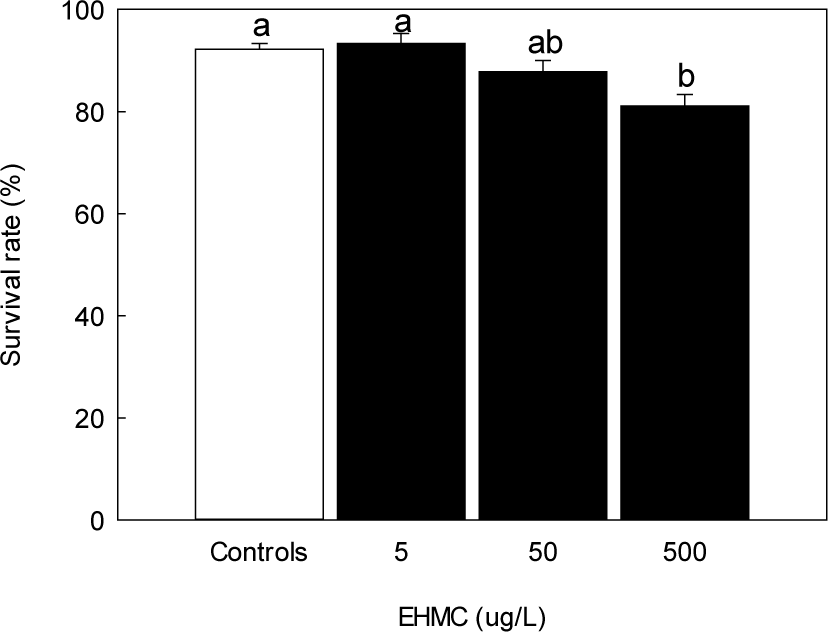
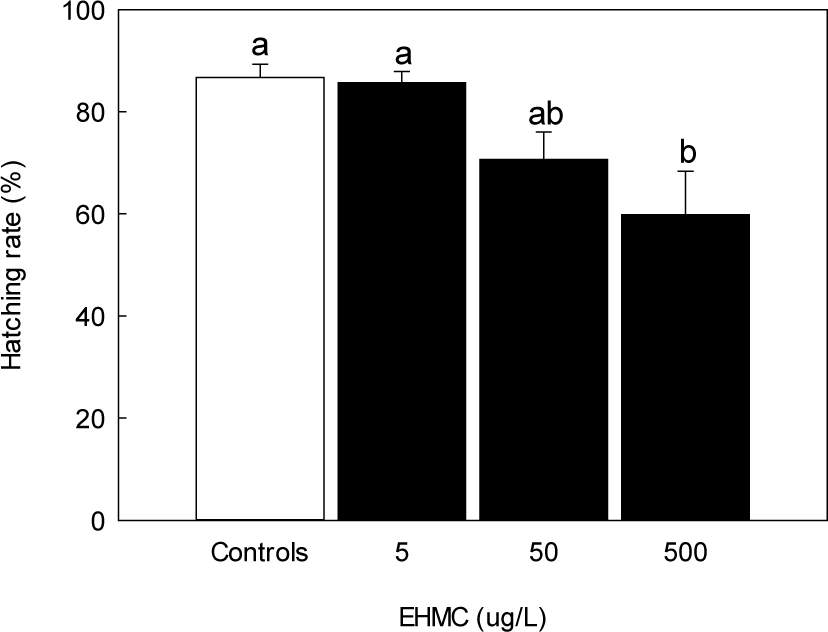
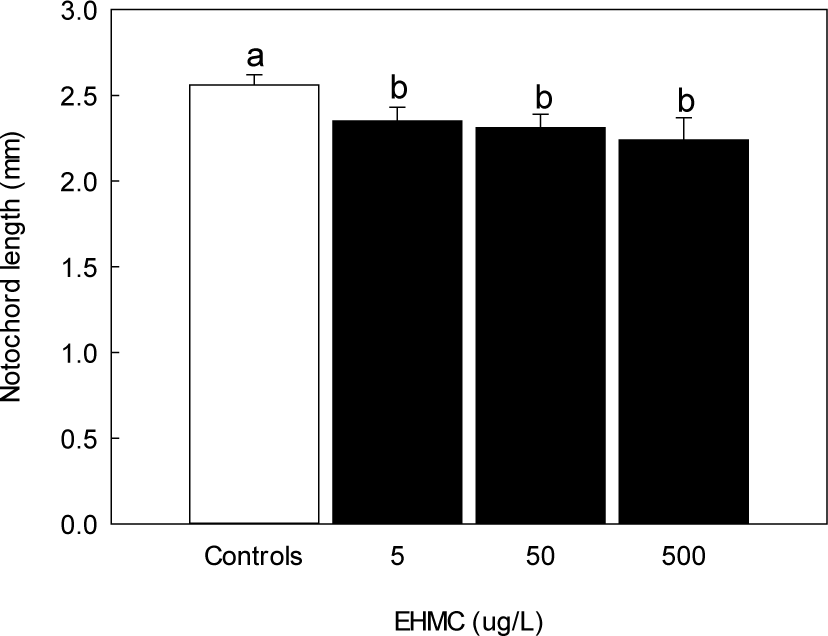
Survival rates at 144 HPE showed statistically significant decrease only at the highest dose, although a slight increase was observed at 5 µg/L, and a decreasing trend occurred at higher concentrations (Fig. 7). In contrast, hatching success declined in a concentration-dependent manner. Hatching rates at 50 and 500 µg/L of EHMC were markedly reduced, with a significant difference observed at 500 µg/L compared to the control (p < 0.05, Fig. 8). The notochord length of newly hatched larvae decreased significantly in all EHMC-treated groups compared to the control, with the greatest reduction observed at 500 µg/L (Fig. 9).
Discussion and Conclusion
The fish embryo acute toxicity test (Organization for Economic Co-operation [OECD] Test Guideline 236) specifies four acute lethality endpoints: coagulation of the embryo, lack of somite formation, non-detachment of the tail, and lack of heartbeat. In addition, various endpoints such as edema formation, BC, and SC are also used as markers for evaluating the toxicity of environmental hazardous substances (von Hellfeld et al., 2020; Wittlerová et al., 2020). In the present study, fertilized eggs of chameleon goby, T. trigonocephalus exposed to EHMC (5, 50, and 500 µg/L) exhibited in a range of adverse effects, including morphological abnormalities, delayed yolk absorption, cardiac dysfunction, and reduced hatching success. Importantly, these effects were observed not only at high exposure concentrations but also at relatively low levels, underscoring the potential ecological risk of EHMC in coastal marine environments. In the previous studies, detected concentrations of EHMC from coastal waters are typically in the range of tens to hundreds of ng/L and rarely approach the low µg/L range although the highest value was 4,043 ng/L in Hong Kong coastal waters (Kim et al., 2017; Langford & Thomas, 2008; Sankoda et al., 2015; Tsui et al., 2014). Therefore, the 5 µg/L of the present study, while exceeding typical ambient monitoring values, represents a conservative upper-range scenario, whereas 50 and 500 µg/L are designated as worst-case or heavily contaminated area (e.g., near-point-source discharge from densely used recreational beaches or cosmetic wastewater effluents).
In the present study, one of the most pronounced effects of EHMC exposure was the induction of morphological abnormalities such as TM and SC. Such structural abnormalities are critical because tail morphology plays a pivotal role in swimming performance, predator avoidance, and feeding efficiency during early life stages (Voesenek et al., 2018). Larvae exhibiting impaired locomotion are more susceptible to predation and starvation, which may ultimately reduce recruitment success in natural populations. Similar tail and spinal deformities have been reported in zebrafish embryos exposed to EHMC and other organic UV filters, suggesting that interference with axial skeletal development may represent a conserved toxicological response across fish species (Yang et al., 2024). In addition, EHMC exposure decreased notochord length of hatched larvae of the present study. Reductions in notochord or body length in hatched larvae represent a functionally significant impairment: shorter larvae are at a competitive disadvantage for predator avoidance, prey capture, rheotaxis, and the energetically critical transition to exogenous feeding (Holley, 2007). Araújo et al. (2018) reported spinal curvature and significantly impaired swimming performance by spinal curvature induction in larvae of the marine flatfish Solea senegalensis following exposure to the UV filter 4-methylbenzylidene camphor (4-MBC), potentially analogous to the structural malformations observed in the present study.
Cardiac dysfunction was another prominent feature of EHMC toxicity observed in the present study. Reduced heartbeat rates, irregular cardiac rhythms, and blood congestion in the pericardial region indicate compromised cardiovascular performance during embryogenesis. The heart is one of the earliest functional organs to develop in vertebrates and is particularly sensitive to chemical exposure (Du et al., 2015; Yalcin et al., 2017). In addition, the reductions in heartbeat rate ranged from approximately 2.8% at 5 µg/L (181.6 vs. 186.8 bpm) to 7.2% at 500 µg/L (173.3 vs. 186.8 bpm) compared to control. While moderate in absolute terms, reductions of this magnitude occurring during critical embryonic developmental windows have been associated with impaired cardiac output, compromised oxygen delivery to developing tissues, and downstream effects on growth and organogenesis (Du et al., 2015; Yalcin et al., 2017). Furthermore, the progressive increase in heartbeat irregularity with increasing EHMC concentration observed in the present study suggests that EHMC may affect both the rate and rhythmicity of cardiac function, which together may carry greater functional significance than rate reduction alone.
Impaired cardiac output may restrict oxygen and nutrient delivery to developing tissues, thereby contributing to growth retardation, developmental delay, and reduced hatching success (Carney et al., 2006). Previous studies have demonstrated that exposure to UV filters alters the expression of genes involved in cardiac development and function, supporting the hypothesis that EHMC may interfere with gene pathways involved in cardiac development, although direct gene expression analysis was not conducted in the present study (Zhao et al., 2023; Zuo et al., 2023). In addition, Nataraj et al. (2020) demonstrated EHMC and its photoproducts induced cardiac abnormalities and developmental delays in zebrafish embryos, supporting the hypothesis that cinnamate-type UV filters share a common cardiotoxic mechanism.
EHMC exposure also significantly delayed yolk sac absorption, indicating disruption of energy utilization during early development. Yolk reserves provide the primary source of energy for embryos and larvae prior to the onset of exogenous feeding (Yu et al., 2019). Delayed or inefficient yolk utilization can reduce energy availability for growth and organogenesis, resulting in smaller body size and impaired physiological performance. Damiani et al. (2023) reported that significant increases in yolk sac area in EHMC-exposed zebrafish larvae relative to controls at 96 hpf, related to delayed yolk lipid mobilization. Previous research has shown that organic UV filters can interfere with lipid metabolism and induce abnormal lipid accumulation in fish embryos, suggesting that EHMC may disrupt embryonic energy homeostasis (Fong et al., 2016). Quintaneiro et al. (2019) reported that zebrafish embryos exposed to 4-MBC showed significantly reduced hatching rates and increased frequencies of tail and yolk sac edema, reinforcing that organic UV filters as a class exert conserved developmental toxicity across fish species.
Survival rate and hatching rate of fertilized eggs are widely used as major endpoints for evaluating early developmental toxicity by chemical exposure (Li et al., 2018; Samaee et al., 2015). Although survival rates were not significantly affected by EHMC exposure, hatching success declined in a dose-dependent manner. This apparent discrepancy suggests that many embryos remained viable but were unable to hatch successfully due to reduced muscular activity or structural malformation.
The slight increase in survival at 5 µg/L EHMC (93.3 ± 3.3%) was not significantly different from control (92.2 ± 1.9%). The reason is unclear, however, we cannot entirely rule out the possibility of a hormetic or low-dose response, a phenomenon that has been documented for some endocrine-disrupting compounds in fish embryo studies (Vandenberg et al., 2012). Future studies with a broader range of low-concentration treatments should be conducted to evaluate this possibility rigorously.
Successful hatching requires both enzymatic degradation of the chorion and sufficient embryonic movement (Korwin-Kossakowski, 2012), processes that may be compromised by EHMC-induced developmental impairments. Similarly, reduced locomotor capacity—including shortened muscle fiber length and decreased responsiveness to external stimuli—as well as delayed hatching have been reported in zebrafish embryos exposed to EHMC (Yang et al., 2024).
Regarding the lowest reported effect, Yang et al. (2024) reported developmental abnormalities in zebrafish embryos at concentrations as low as 100 µg/L (EHMC), while Zhou et al. (2019a) found increased malformation and mortality at 50 µg/L with parental exposure. Damiani et al. (2023) observed significant yolk sac retention in zebrafish more than 10 µg/L. In the present study, TM were observed at 5 µg/L, heartbeat reductions were significant across all treatments (5–500 µg/L), and notochord shortening was detected at 5 µg/L—indicating that the chameleon goby embryo may be comparably or more sensitive than zebrafish at the lowest concentration of EHMC. We note that direct comparison with marine fish species is limited by the scarcity of published studies, further underscoring the value of the present study.
Several limitations of the present study should be acknowledged. First, measured concentrations of EHMC in the incubation media were not analytically verified. The exposure concentrations of the present study were nominal values. Due to the hydrophobic nature of EHMC, adsorption to the walls of 6-well polystyrene plates may have resulted in actual dissolved concentrations lower than nominal values, particularly at higher treatment levels. Consequently, concentration–response relationships and effect thresholds reported herein should be interpreted as conservative estimates based on nominal concentrations, and future studies should incorporate analytical verification of actual exposure concentrations using high-performance liquid chromatography (HPLC) or liquid chromatography-tandem mass spectrometry (LC-MS/MS) methods. Second, while morphological and physiological endpoints provided valuable insights, integration of molecular and biochemical markers would allow for a more comprehensive understanding of the mechanisms underlying EHMC toxicity. Finally, long-term post-hatching studies are needed to determine whether early-life exposure to EHMC results in persistent effects on growth, behavior, and reproductive capacity.
In addition to elucidating the developmental toxicity of EHMC, this study highlights the utility of the chameleon goby as a marine model species for ecotoxicological research. This species readily spawns under laboratory conditions, exhibits well-defined embryonic and larval developmental stages, and responds sensitively to chemical exposure. Given the limitations of extrapolating freshwater toxicity data to marine ecosystems, the use of marine-relevant model species is essential for accurate ecological risk assessment. The embryo toxicity assessment model for chameleon goby proposed in this study is expected to serve as foundational data not only for evaluating the effects of EHMC but also for assessing the impacts of various other environmental hazardous substances.
In conclusion, the present study demonstrates that EHMC poses a developmental hazard to marine fish by interfering with key physiological and morphological processes during early life stages. These findings contribute important marine-specific toxicity data and emphasize the need for regulatory frameworks that incorporate sensitive early-life-stage endpoints and marine species in the environmental risk assessment of UV filters.