
Introduction
Aquaculture is one of the fastest-growing sectors of global agro-production industry, with Asia alone accounting for over 70% of total farmed fish output (Taslima et al., 2024). Beyond its economic importance, aquaculture also plays a key role in food and nutritional security by providing a stable supply of high-quality proteins, essential fatty acids, vitamins, and minerals (Verdegem et al., 2023). This rapid expansion is largely driven by increasing demand for aquatic products and declining wild fish stocks, leading to a growing focus on the cultivation of high-value marine species (Little et al., 2016). In Southeast Asia, Vietnam endowed with rich bioresources across its marine and coastal ecosystems has emerged as a major aquaculture producer, supported by its extensive coastline exceeding 3,260 km (Giang & Khanal, 2024). The Vietnamese waters host more than 2,000 documented marine fish species, of which a limited subset is of commercial relevance to capture fisheries and aquaculture (Tran et al., 2016). Among cultivated marine fishes, species of the genus Trachinotus Lacépède, 1801 (order Carangiformes; family Carangidae; subfamily Trachinotinae), commonly known as pompano, are widely farmed due to their rapid growth, tolerance to farming conditions, high market value, and consistent consumer demand (Chen et al., 2024). The commercial mariculture of pompano in Asia began in the early 1990s and has since expanded substantially, contributing to a current global production over 110,000 tons annually (FAO, 2026). Two main species are cultured: the long fin-clip pompano (Trachinotus blochii) (Nguyen et al., 2025) and the short fin-clip pompano, which has been reported under three different scientific names (Trachinotus ovatus, Trachinotus anak, and Trachinotus falcatus).
Despite its considerable commercial value, the identity of Trachinotus species used in Asian mariculture remains uncertain (Smith-Vaniz & Walsh, 2019). Species identification of Trachinotusin aquaculture has traditionally relied on external morphological traits; however, such characters are often unreliable due to phenotypic plasticity and overlapping features among closely related species (Shadrin et al., 2024). A prominent taxonomic controversy involves the frequent reference to T. ovatus (Linnaeus 1758) as a commonly farmed species in Vietnam and China, despite its native range being restricted to the Eastern Atlantic and Mediterranean regions (Fricke et al., 2026; Smith-Vaniz, 1986). Notably, there is no scientific evidence supporting the natural occurrence or intentional introduction of T. ovatus into Asian marine ecosystems, including Vietnamese waters (Shadrin et al., 2024; Welch, 2013). A recent integrative study combining morphological analyses with DNA barcoding of pompano specimens collected from aquaculture systems in central Vietnam preliminarily indicated the presence of only two Trachinotus species, T. anak Ogilby 1909 and T. blochii (Lacepède 1801) (Shadrin et al., 2024). The study further reported that T. anak is the dominant species cultured in aquaculture systems and is frequently misidentified as T. falcatus (Linnaeus 1758) or T. ovatus. These findings strongly suggest that widespread species misidentification persists in aquaculture operations and commercial labeling across Vietnam and neighboring countries. However, the previous research utilized only a limited number of T. anak congeners (T. ovatus, Trachinotus baillonii, T. blochii, T. falcatus, and Trachinotus mookalee), which is insufficient to fully resolve phylogenetic relationships, accurately discriminate among species, or elucidate population genetic structure (Shadrin et al., 2024). Therefore, given the broad applicability and significance of molecular tools in biodiversity and systematic research, a more comprehensive assessment is required to validate and extend earlier investigation (Shadrin et al., 2024).
In this context, genetic tools targeting the mitochondrial cytochrome c oxidase subunit I (COI) gene are widely recognized as reliable markers for accurate species identification. Their robustness is based on estimates of intra- and inter-specific genetic variation, analyses of population genetic structure, and the application of monophyly criteria through phylogenetic inference (Appleyard et al., 2025; Ewusi et al., 2025). Nevertheless, genetic identification alone does not capture other biological attributes related to physiological condition, which are also critical for effective aquaculture management. Accordingly, the length–weight relationship (LWR) represents a fundamental biological parameter for evaluating growth patterns, stock structure, and ecological adaptation in cultured fish populations (Biswas et al., 2011). Although fish weight is generally expected to scale with the cube of body length, deviations from this relationship may occur across different ontogenetic stages because growth coefficients are influenced by culture-specific factors, such as feeding management and stocking density (Mazumder et al., 2016). Thus, estimating LWR for a valid species in mariculture systems enables the assessment of biological performance and population condition, thereby supporting more informed and sustainable aquaculture management (Bordoloi et al., 2024). Hence, the present study investigates morphologically identified as T. anak from an aquaculture site in northern Vietnam through genetic analyses and LWR assessments. Specifically, the study aims to (i) construct phylogenetic relationships to validate species identity of T. anak; (ii) estimate molecular operational taxonomic units (MOTUs) using multiple species-delimitation methods and assess genetic distinctiveness among different pompano species; (iii) examine the inter-species haplotype diversity (Hd) to reevaluate the genetic differentiation between T. anak and T. ovatus, as well as their population structure; and (iv) evaluate LWR analyses as indicators of population condition and wellbeing in cultured conditions. Overall, this study provides a clearer understanding of the species identity and genetic diversity of T. anak from a single mariculture site in northern Vietnam in comparison with global populations of T. anak and T. ovatus. These findings establish a preliminary scientific foundation to support the sustainable development and management of pompano mariculture in Vietnam and other Asian countries.
Materials and Methods
Initially, a total of 327 pompano specimens of similar age (300 days) were collected in March 2025 from an aquaculture site in Cam Pha City, Quang Ninh Province, Vietnam (20.9849° N, 107.2657° E) (Fig. 1B) for morphological examination and LWR analysis. The species was subsequently identified as T. anak based on diagnostic morphological characters, following previous literatures (Smith-Vaniz & Walsh, 2019; Shadrin et al., 2024). Notably, the species displays a dorsal coloration ranging from silvery-green to bluish-gray, gradually transitioning to a lighter ventral surface. The body often exhibits a bronze or golden-green iridescence, while the second dorsal and caudal fins vary in color from dull orange to nearly black, typically with darker anterior edges. The anal fin is bright yellow, resembling the hue of dandelion petals; the pelvic fins are paler, and the pectoral fins show relatively darker pigmentation (Fig. 1A). The body weight was measured for each specimen using an electronic balance with 0.01 g precision, and total length was recorded to the nearest 0.1 cm using a standard measuring board. Additionally, 24 specimens were randomly selected for downstream molecular analysis. Approximately 30 mg of muscle tissue was aseptically excised from the lateral line of each specimen, transferred into 2 mL centrifuge tubes containing 95% molecular-grade ethanol, and stored at –20°C. In addition, the research and experimental procedures have followed the standards for animal research in Vietnam from 8/2024 to 8/2027 via approval decision number 19/2024/106/HĐTN, and all procedures were conducted in accordance with the ARRIVE 2.0 guidelines (https://arriveguidelines.org/) (Percie du Sert et al., 2020).
The genomic DNA was extracted using the AccuPrep®Genomic DNA Extraction Kit (Bioneer, Daejeon, Korea). Each muscle tissue sample was mixed with 600 μL of 1× lysis buffer and homogenized for 60 seconds using a TissueLyser II (Qiagen, Hilden, Germany). To enhance cell lysis and protein degradation, 100 μL of sodium dodecyl sulfate (SDS) and 20 μL of proteinase K were added to the homogenate, followed by incubation at 60°Cfor 12 hours. The DNA precipitation was carried out by adding 500 μL of GC buffer and 300 μL of isopropanol. The mixture was subsequently transferred to a spin column and centrifuged at 8,000 rpm for 1 minute. The column-bound DNA was washed sequentially with Wash Buffer 1 and Wash Buffer 2 to eliminate residual contaminants, and the purified DNA was eluted in 50 μL of TE buffer. The DNA concentration and purity were assessed using a NanoDrop spectrophotometer (D1000, Thermo Fisher Scientific, Waltham, MA, USA).
To amplify the partial mitochondrial COI gene fragment, the universal primers Fish-BCH (5'-TCAACYAATCAYA AAGATATYGGCAC-3') and Fish-BCL (5'-ACTTCYGGGTGR CCRAARAATCA-3') were used in this study (Baldwin et al., 2009). Each polymerase chain reaction (PCR) (30 μL total volume) consisted of 1 μL of each primer, 0.9 μL of 3% dimethyl sulfoxide (DMSO), 19.9 μL of nuclease-free water, 3 μL of 10× Ex Taq buffer, 0.2 μL of Ex Taq Hot Start DNA polymerase, 3 μL of dNTP mix, and 1 μL of 1:10 diluted genomic DNA. The thermal cycling protocol included an initial denaturation at 94°Cfor 3 minutes, followed by 40 cycles of denaturation at 94°Cfor 30 seconds, annealing at 50°Cfor 30 seconds, and extension at 72°Cfor 1 minute, with a final extension at 72°Cfor 5 minutes. The PCR products were electrophoresed on a 1.5% agarose gel stained with ethidium bromide alongside a 1 kb DNA ladder and visualized under ultra violiet (UV) illumination. The successfully amplified products were then purified using the AccuPrep®PCR/Gel Purification Kit (Bioneer, Daejeon, Korea) and sequenced bidirectionally using an ABI PRISM 3730XL DNA Analyzer (Macrogen, Daejeon, Korea). The chromatograms were visually inspected and edited using SeqScanner v1.0 (Applied Biosystems, Foster City, CA, USA) to remove ambiguous bases and low-quality regions. The final sequences of T. anak were deposited in the GenBank database for acquiring unique accession numbers.
A comprehensive dataset was compiled, consisting of 24 newly generated COI sequences and 443 additional sequences retrieved from GenBank, representing 17 Trachinotus species (Supplementary Table S1). The sequence alignment was conducted using CLUSTAL X (Thompson et al., 1997), yielding a consensus alignment of 620 bp. The COI sequence of Lichia amia (Carangidae: Trachinotinae; GenBank Accession No. JQ623944) was used as the outgroup for phylogenetic analyses (Keskin & Atar, 2013). The phylogenetic trees were reconstructed using both Bayesian inference (BI) and maximum-likelihood (ML) approaches. The best-fit nucleotide substitution model, General Time Reversible with gamma-distributed rate heterogeneity and a proportion of invariant sites (GTR+G+I), was selected according to the lowest Bayesian information criterion (BIC) scores, as identified by PartitionFinder 2 (Lanfear et al., 2017). The BI analysis was performed in MrBayes v3.1.2 using the nst = 6, employing a Markov chain Monte Carlo (MCMC) algorithm run for 1,000,000 generations with four chains (one cold and three heated). The sampling was conducted every 100 generations, and the initial 25% of trees were discarded as burn-in (Ronquist et al., 2012). The ML analysis was carried out using the PhyML 3.0 web server (http://www.atgc-montpellier.fr/phyml/), with 1,000 bootstrap replicates to assess the robustness of the inferred clades (Guindon et al., 2010). The resulting phylogenetic trees from both BI and ML analyses were visualized using the Interactive Tree of Life (iTOL) platform v4 (https://itol.embl.de/) (Letunic & Bork, 2024).
To robustly delineate MOTUs among 17 Trachinotus species, an integrative approach incorporating multiple molecular delimitation methods was applied. These included automatic barcode gap discovery (ABGD), assemble species by automatic partitioning (ASAP), and poisson tree processes (PTP) models. Notably, the sequence alignments in FASTA format were analyzed using ABGD and ASAP under the Jukes-Cantor (JC69) substitution model (Puillandre et al., 2012, 2021). Differently, the PTP required an ultrametric phylogenetic tree generated via the ML method and formatted in Newick (.nwk) to estimate the number of putative MOTUs (Fujisawa & Barraclough, 2013; Zhang et al., 2013). The species delimitation analyses for ABGD and ASAP were carried out through the Species PARTition (SPART) online platform, whereas the PTP was executed using the iTaxoTools v0.1 (Miralles et al., 2022; Vences et al., 2021). The genetic distances within and between species were calculated based on the Kimura 2-Parameter (K2P) model implemented in MEGA v12 (Kumar et al., 2024).
The population genetic structure of 241 partial mitochondrial COI gene sequences labeled as T. anak and T. ovatus, obtained from both newly generated data and global GenBank records was analyzed. These datasets were subsequently used to estimate the number of haplotypes, Hd, and nucleotide diversity (π) using DnaSP v6.0 (Rozas et al., 2017). The haplotype relationships were visualized using a Templeton, Crandall, and Sing (TCS) network constructed in POPART v1.7 (Clement et al., 2000; Leigh & Bryant, 2015). Subsequently, based on the results obtained from the haplotype network analysis, the selective neutrality tests were further conducted at each locality using Tajima’s D, as well as Fu and Li’s D and F statistics. These analyses were also performed using DnaSP v6.0, based on sequence data in FASTA format and encompassing phylogenetically specified clades of T. anak and T. ovatus. The calculations employed a sliding window approach with a length of 100 nucleotide sites and a step size of 25 sites (Fu & Li, 1993; Tajima, 1989).
To provide baseline LWR information for T. anak under site-specific study conditions, a total of 327 specimens, ranging in length from 21 to 40.9 cm, were collected from a single mariculture site during a single sampling period for analysis. The total length (cm) of all specimens was measured from tip of snout to longest axis of caudal fin, through measuring board. The body weight (g) of each individual fish was also recorded along with the corresponding length. The specimens fall in the range from 234.9 to 1257.2 g. The LWR of the studied fish species was estimated using the standard power function W = aLb, where W represents the observed body weight (g), L is the measured total length (cm), a is the intercept of the relationship, and b is the allometric growth exponent (López-Pérez et al., 2020; Ragheb, 2023). To estimate the parameters a and b, the equation was linearized using logarithmic transformation, resulting in the linear form: log10(W) = log10(a) + b log10(L). The parameters were estimated using ordinary least-squares regression of log10(W) on log10(L). The slope (b) was calculated using the least-squares formula: . The intercept was calculated as . The parameter a was obtained by back-transforming the intercept of the regression a = 10log10(a). To determine whether growth was isometric or allometric, the null hypothesis of isometric growth (H0: b= 3) was tested using t-test as t = (b − 3) / SE(b), where SE(b) is the standard error of the slope and the degrees of freedom were N − 2. The expected weight of each individual fish was calculated using the estimated LWR equation , and the relative condition factor was computed as , where W is the observed weight and is the predicted weight derived from the LWR model.
Results
The morphological examination of pompano specimens collected from a mariculture site in northern Vietnam preliminarily identified them as T. anak based on diagnostic body and fin coloration patterns. To validate this morphological identification, the molecular approach was further implemented using partial sequences of the mitochondrial COI gene. The nucleotide Basic Local Alignment Search Tool (BLAST) search of the generated sequences revealed a 99.6%–100% top similarity hits with T. ovatus originating from China. This result raises concerns, as the historical geographic distribution of T. ovatus has been primarily confined to the Eastern Atlantic and the Mediterranean Sea. Thus, to confirm species-level identification, a combination of phylogenetic inference, estimation of MOTUs, inter- and intra-specific genetic diversity analyses, and haplotype network construction was performed using both newly generated and GenBank-derived sequences of T. ovatus and T. anak, following verification of their collection localities from prior studies and database records. In addition, the comparative molecular analyses incorporating 15 other valid congeners further substantiated the species delineation and genetic distinctiveness of both T. ovatus and T. anak.

Based on the current dataset, phylogenetic analyses using both BI and ML methods revealed that most of the Trachinotus showed clear-cut clustering with high posterior probability and bootstrap supports, except Trachinotus goreensis and Trachinotus teraia (Fig. 2 and Supplementary Fig. S1). The 30 database sequences labeled as T. ovatus, originating from Morocco, Türkiye, Angola, Tunisia, Ghana, and Portugal, consistently formed a cohesive cluster (Clade-1) in both BI and ML phylogenetic trees. Conversely, the 24 sequences generated in the present study, along with 187 database sequences labeled as either T. ovatus or T. anak sourced from China, Vietnam, Australia, and the United Kingdom (international trade) clustered separately as a distinct lineage (Clade-2), indicating a clear cladding pattern from the real T. ovatus clade. The phylogenetic tree topologies also demonstrated a close genetic relationship between T. anak, which is distributed across Eastern and Northern Australia, southern part of the China Sea, and Southern Japan, and T. mookalee, distributed in the Persian Gulf and Gulf of Oman eastward to Eastern China (East China Sea), and northward to Southern Japan. In contrast, T. ovatus, which occurs in the Western Baltic Sea, North Sea, Mediterranean Sea, Sea of Marmara, Black Sea, and across the Eastern and Southern-Central Atlantic, was found to be genetically more closely related to Trachinotus botla, a species native to the Eastern Cape and KwaZulu-Natal (South Africa), East Africa, Socotra (Yemen), and Madagascar, extending eastward to the eastern coast of India, Myanmar, and Western Australia (Fig. 2 and Supplementary Fig. S1).
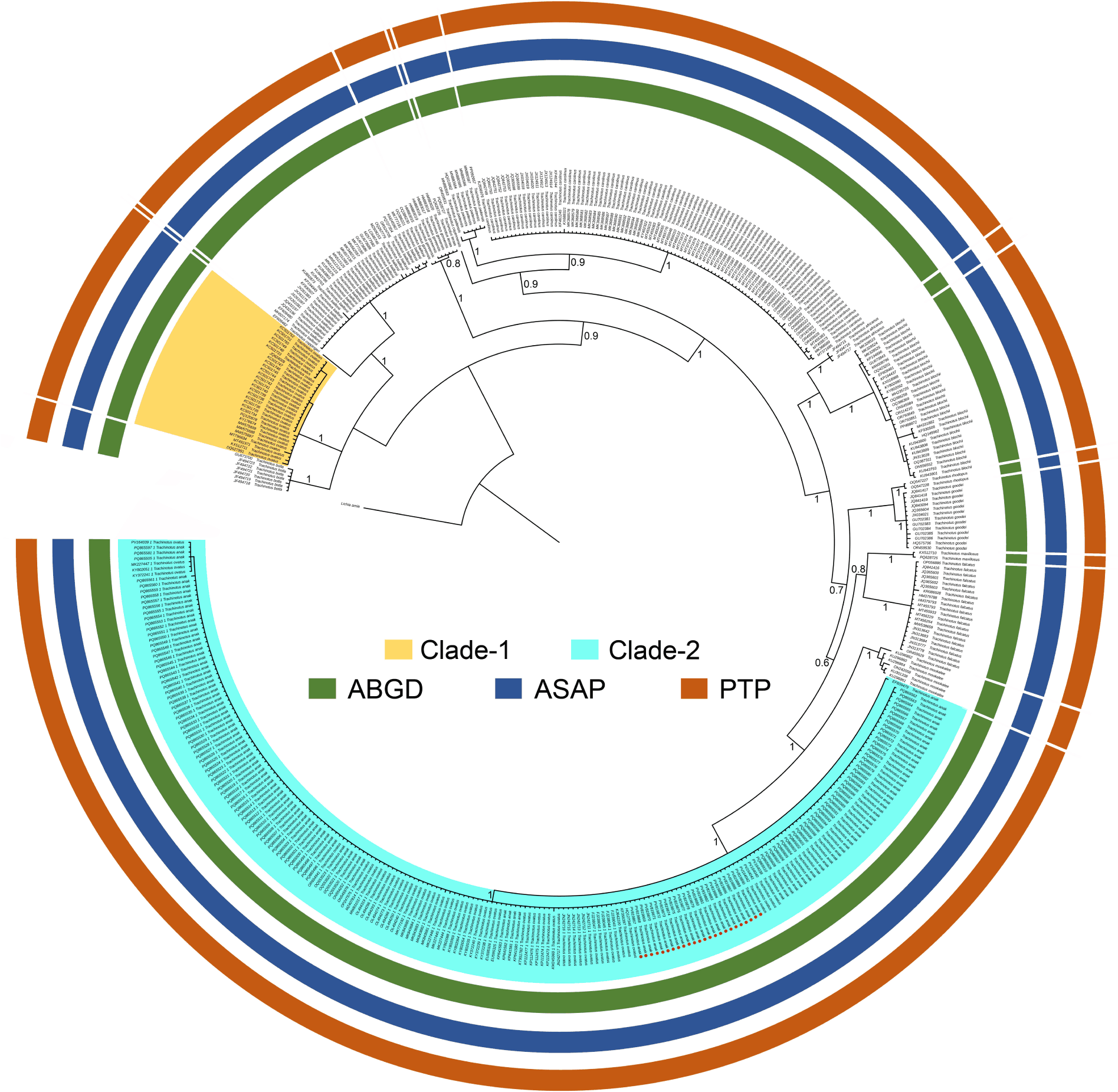
Furthermore, three species delimitation approaches consistently identified 16 distinct MOTUs within the dataset comprising 17 Trachinotus species. Notably, Clade 1 (T. ovatus) and Clade 2 (T. ovatus + T. anak) were each delineated as separate MOTUs, while T. goreensis and T. teraia were consistently grouped within a single MOTU across all methods, indicating potential taxonomic ambiguity requiring further systematic revision (Fig. 2 and Supplementary Table S2). Overall, both the phylogenetic analyses and MOTU estimations confirmed the species identities of T. ovatus (Clade-1) and T. anak (Clade-2) by reinforcing their genetic distinctiveness and distinct clustering patterns. In addition, the genetic divergence based on the K2P model using partial COI sequences ranged from 0.0% to 18.4%. The lowest mean inter-specific genetic divergence (0.0%) was detected between T. goreensis and T. teraia, whereas the highest divergence (18.4%) was observed between T. mookalee and T. baillonii. Notably, the genetic distance between T. anak and T. ovatus was 15.2%, indicating substantial inter-specific differentiation. The studied species, T. anak exhibited the lowest genetic distance with T. mookalee (5.5%) and the highest with T. baillonii (17.4%). In terms of mean intra-specific variation, T. anak displayed the lowest divergence (0.1%), while the highest intra-specific divergence was recorded in T. maxillosus (5.7%) (Table 1).
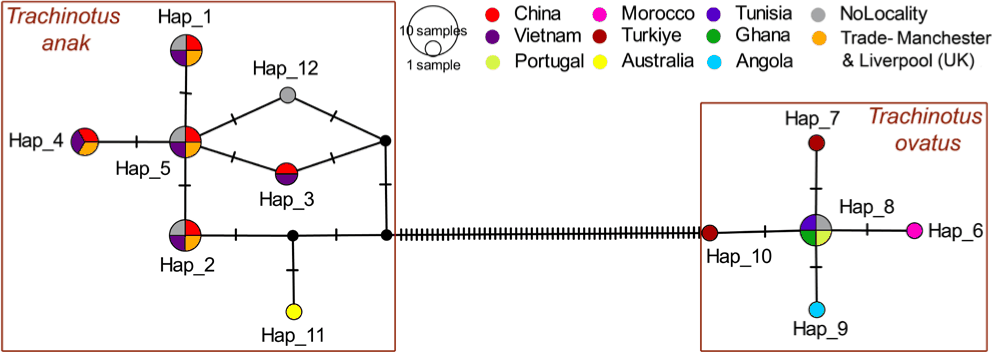
The haplotype network analysis of T. anak and T. ovatus sequences clearly illustrated distinct clustering between these two species, reflecting significant genetic differentiation. The combined dataset for T. anak and T. ovatus revealed a Hd = 0.5315, π = 0.03096, and identified 12 distinct haplotypes (Fig. 3 and Supplementary Table S3). The haplotype network highlighted two distinct clusters corresponding to T. anak and T. ovatus, with a high level of genetic variation reflected by 66 polymorphic sites. Notably, the cluster of valid T. anak exhibited seven haplotypes, most of which were shared among individuals from China, Vietnam, and the United Kingdom (international trade). Further, within the T. anak cluster, one unique haplotype was identified from Australia, and another from a sample lacking locality information. Conversely, the T. ovatus displayed five haplotypes, including four unique ones, Hap_6 (Morocco), Hap_7 and Hap_10 (Türkiye), and Hap_9 (Angola), while a single shared haplotype (Hap_8) occurred in samples from Tunisia, Ghana, Portugal, and sequences lacking locality information (Fig. 3 and Supplementary Table S3). Moreover, in T. anak, eight polymorphic sites and seven haplotypes were identified, with an average number of nucleotide differences (K) = 0.46572, Hd = 0.3986, and π = 0.00095. In contrast, the T. ovatus presented four polymorphic sites and five haplotypes, with K = 0.63448, Hd = 0.4828, and π = 0.00136. Interestingly, the 24 sequences of T. anak generated in this study exhibited considerable genetic variation, encompassing three haplotypes (Hap_2, Hap_3, and Hap_5). Specifically, two sequences (PV819885 and PV819887) clustered within Hap_2 along with sequences generated from China and the United Kingdom, likely reflecting patterns associated with international trade. In contrast, Hap_3 comprised three sequences (PV819876, PV819881, and PV819888), which were grouped with sequences originating from China that shared the same haplotype. Furthermore, the remaining 19 sequences were assigned to Hap_5 and clustered with sequences from China and the United Kingdom (Supplementary Table S3). Additionally, the neutrality test results indicated that T. anak exhibited Tajima’s D = –1.43442, Fu and Li’s F = –2.36381, and Fu and Li’s D = –2.34870, whereas T. ovatus demonstrated Tajima’s D = –0.95412, Fu and Li’s F = –1.81305, and Fu and Li’s D = –1.98083 (Table 2).
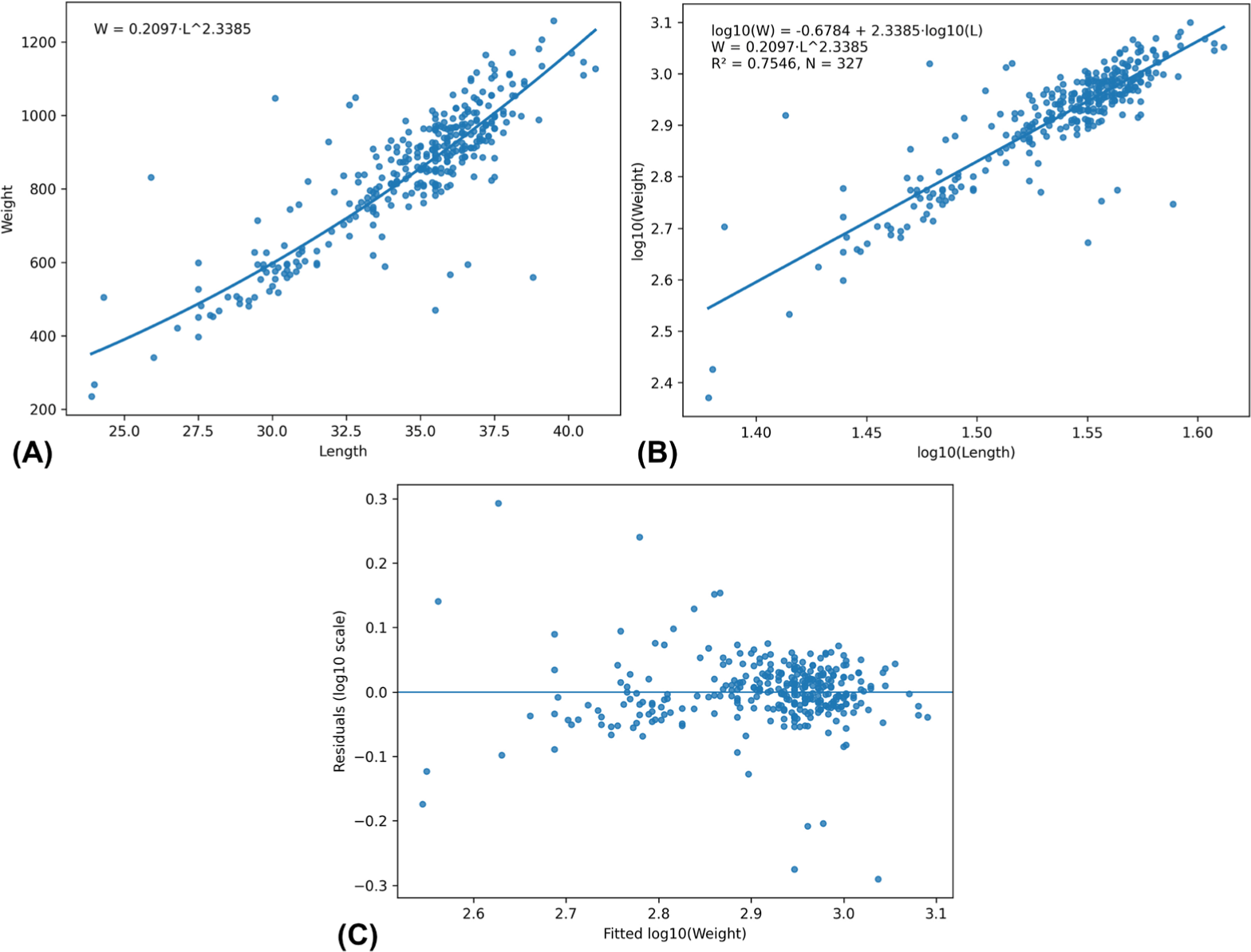
A total of 327 specimens were analyzed to estimate the LWR of the studied fish species. The relationship between total length and body weight showed a significant positive correlation and was best described by the power function W = 0.2097 L2.3385 (Fig. 4A). After logarithmic transformation, the regression equation was expressed as with a coefficient of determination of R2 = 0.7546 (Fig. 4B and Supplementary Table S4), indicating that approximately 75.46% of the variation in body weight was explained by variation in total length. The estimated exponent b = 2.3385 was significantly lower than the standard value of 3, which represents isometric growth. The t-test confirmed that the difference was statistically significant (t = −8.9419, df = 325, p < 0.001), and the 95% confidence interval for b ranged from 2.1930 to 2.4840 (Supplementary Table S4). These results indicate that the species exhibits negative allometric growth, meaning that body weight increases at a slower rate relative to increases in length. The predicted weights calculated from the LWR model were used to estimate the relative condition factor (Kn) for each individual fish (Supplementary Table S5), which provides an index of the physiological condition of the population relative to the expected weight at a given length (Fig. 4C).
Discussion
Morphological characteristics such as variations in fin morphology, body coloration, and pigmentation patterns are often recognized as important diagnostic traits for distinguishing congeners within the genus Trachinotus (pompano). The morphological assessment conducted in the present study indicates that T. anak is characterized by a silvery-green to bluish-gray dorsal coloration that gradually transitions to a lighter ventral surface, accompanied by a bronze to golden-green iridescence across the body. The species also displays distinctive fin coloration, including dull orange to nearly black second dorsal and caudal fins, a bright yellow anal fin, paler pelvic fins, and comparatively darker pectoral fins. In contrast, previous descriptions indicate that T. ovatus exhibits a greenish-gray dorsal surface and silvery flanks marked by 3–5 vertically elongated black spots along the anterior portion of the lateral line, while the lobes of the dorsal, anal, and caudal fins are typically tipped with black (Bauchot, 2003). Based on these morphological variations, the two species can be readily distinguished; however, the integration of molecular data is recommended as a complementary approach to strengthen and validate this taxonomic delimitation.
In recent years, biodiversity and systematics research has accelerated rapidly, driven by the exponential growth of molecular data and its widespread open access through global databases (Rouard et al., 2025). Among them, the GenBank database provides an excellent platform for benefit-sharing reference digital sequence information (DSI), which is extensively utilized through BLAST-based sequence comparisons (Armani et al., 2015; Blanco-Fernandez et al., 2023). Thus, all generated DSI should be deposited with meticulous care by data contributors and rigorously curated by database managers to ensure accuracy and reliability (Leray et al., 2019). Similarly, end users must exercise caution to minimize the risk of mislabeling or misidentification of reference sequences, as such errors can propagate taxonomic and systematic inaccuracies (Díaz-Arce & Rodríguez-Ezpeleta, 2023). These issues are particularly frequent among marine species, such as pompano, which exhibit high morphological similarity and broad geographic distributions, potentially undermining the validity of scientific findings (Blanco-Fernandez et al., 2021). Notably, the misidentification problem is exemplified by numerous studies from China reporting the presence of T. ovatus as a local population based on genetic analyses and present BLAST search findings (Shadrin et al., 2024). However, definitive information regarding the timing and pathways of introduction of this valid species from its native range in the Eastern Atlantic and Mediterranean Sea remains questionable, as neither biogeographic literature nor historical records provide a comprehensive explanation of its initial occurrence in Asian waters (Shadrin et al., 2024; Welch, 2013). In this context, the present study employs various molecular analyses to investigate and validate the recent finding of misidentification between T. ovatus and T. anak (Smith-Vaniz & Walsh, 2019; Shadrin et al., 2024). The misidentification between these species has profound implications, affecting biodiversity assessments, systematic research, and the sustainability of aquaculture by distorting ecological, genetic, and conservation evaluations (Vindigni et al., 2021). From a practical perspective, such discrepancies also hinder selective breeding programs and compromise effective broodstock management, particularly in aquaculture practices (Garcia-Vazquez et al., 2012).
More specifically, the phylogenetic analyses, MOTUs, and haplotype networks reveal significant inconsistencies between sequences labeled as T. ovatus in GenBank and the results of current molecular assessments. The GenBank records labeled T. ovatus from China, Vietnam, and the United Kingdom (via international trade) clustered within Clade-2 in association with T. anak. Conversely, the taxonomically validated T. ovatus sequences from Morocco, Türkiye, Angola, Tunisia, Ghana, and Portugal form a distinct Clade-1. These findings are further supported by previous scientific investigation, which revealed that specimens marketed as T. ovatus in fish trades in Manchester and Liverpool (UK) were, in fact, sourced from Vietnam (Di Muri et al., 2018). Moreover, the high level of genetic distance (15.2%) between T. ovatus and T. anak substantiates their classification as distinct species. In the particular case of T. anak, although all 24 samples were collected from the single aquaculture facility in northern Vietnam, the presence of three haplotypes in this study indicates existing genetic variation within the cultured stock. This pattern may reflect the use of multiple broodstock sources or the introduction of individuals originating from different geographic populations through aquaculture trade. Interestingly, database sequences of two other Trachinotus species, T. goreensis (accession nos. HM883015–HM883020) and T. teraia (accession nos. PQ628726–PQ628727), exhibited 0% mean inter-specific genetic distance and clustered within the same clade in the phylogenetic analysis, forming a single MOTU. This pattern suggests a potential misidentification between these two nominally valid species, which may be associated with their highly similar and overlapping native biogeographic distributions. Specifically, T. goreensis has been reported from the eastern Atlantic, ranging from Mauritania south to Angola, including the Cape Verde Islands and São Tomé and Príncipe, whereas T. teraia occurs in the eastern Atlantic from Senegal south to Angola, also encompassing the Cape Verde Islands and São Tomé and Príncipe (Fricke et al., 2026). In line with these findings, we emphasize the critical state of global molecular databases, which continue to harbor numerous misidentified DNA sequences due to inaccurate morphological-based species identifications. This issue is particularly prevalent in Southeast and East Asian mariculture, where unresolved taxonomic ambiguities undermine the reliability of downstream scientific research, evidence-based aquaculture practices, and effective conservation planning (Naz et al., 2023). To address this, rigorous validation of genomic data prior to submission to public repositories is essential to ensure taxonomic accuracy, uphold scientific integrity, and prevent misleading conclusions in biodiversity research (Shadrin et al., 2024). The outcomes of the present study corroborate earlier investigation and resolve the longstanding taxonomic dilemma between T. anak and T. ovatus in Vietnam and beyond.
Furthermore, based on the dataset of this study, both T. anak and T. ovatus exhibited negative values in various neutrality tests, (Tajima’s D, Fu and Li’s D, and Fu and Li’s F). However, the statistical significance was not detected (p > 0.05), therefore these values do not provide sufficient evidence to infer recent population expansion or purifying selection. The negative neutrality statistics alone may indicate a trend toward an excess of rare alleles, but such interpretations require statistically significant results (Fu & Li, 1993; Tajima, 1989). Such signals are particularly relevant in mariculture contexts, as they may reflect selective pressures favoring traits associated with growth, survivability, or adaptation to captive environments (Uusi-Heikkilä, 2020). These patterns might also be influenced by founder effects due to the introduction of limited wild broodstock or the integration of individuals from different genetic traits to enhance culture functioning (Ansah et al., 2014). The neutrality tests further validate the distinct genetic differentiation between T. anak and T. ovatus, reinforcing their separation as independent biological entities. This distinction is crucial in mariculture, as it underscores the importance of species-specific genetic monitoring (Skibinski, 2000).
The estimated LWR for T. anak in the present study followed the power model W = 0.2097L2.3385, indicating a negative allometric growth pattern (b < 3). The estimated exponent b = 2.3385 was significantly lower and confirming that body weight increased at a slower rate relative to length. Such negative allometric growth suggests that individuals become relatively more elongated or lighter as they increase in size, a pattern commonly reported in fish populations subjected to environmental or husbandry influences. The R2 observed here (~0.75) suggests moderate-to-strong length–weight coupling, while remaining scatter may reflect natural variability (e.g., condition, maturity stage, sex, season, feeding status, or sampling heterogeneity), which can be explored further by stratified LWRs if such metadata are available. Nevertheless, the growth patterns in fish are known to be affected by several biological and environmental factors, including food availability, water quality, stocking density, temperature, and culture management practices. Under mariculture conditions, such factors may influence energy allocation between somatic growth, maintenance, and reproduction, potentially resulting in deviations from isometric growth (Komba et al., 2020). Similar growth patterns have been reported in cultured fish populations under site-specific environmental conditions (Chandran et al., 2023). Because fish growth is highly responsive to local environmental conditions and aquaculture practices, the present findings emphasize the importance of conducting localized assessments of growth performance in cultured stocks. Therefore, this study provides important baseline information on the length–weight scaling and physiological condition of T. anak cultured under mariculture conditions in northern Vietnam. Furthermore, the observed variability in body weight among individuals of similar lengths may partly reflect biological factors such as reproductive status, where gonadal development can influence somatic weight, particularly in mature females (Wagaw et al., 2024).
Limitations and Recommendations for Future Research
Although this study confirms the presence of T. anak in the mariculture system of northern Vietnam through clear genetic differentiation from T. ovatus, and corroborates earlier report of misidentification between these species, it also provides preliminary LWR data relevant to aquaculture management. However, several limitations should be considered. Specifically, the taxonomic coverage was incomplete, as molecular analyses included only 17 of the 21 globally recognized valid Trachinotus species, which is insufficient for robust phylogenetic construction. Further, the reliance on a single matrilineal marker constrains phylogenetic resolution, particularly for resolving deeper cladistic relationships within the genus. In addition, both molecular and LWR data for T. anak were derived exclusively from cultured specimens from a single aquaculture facility, which may not adequately represent natural genetic variation or growth dynamics observed in wild populations. The artificial and relatively homogeneous rearing conditions, including stable water quality, uniform feeding regimes, constant stocking densities, and limited habitat heterogeneity may influence both growth parameters and genetic structure. To address these limitations, future studies should prioritize expanding taxonomic sampling to include all valid Trachinotus species and adopt multi-locus molecular approaches integrating both mitochondrial and nuclear markers to enhance phylogenetic resolution and population genetic structure. Nevertheless, incorporating samples from both wild and cultured populations of T. anak would facilitate comparative assessments of genetic diversity and growth patterns, thereby enhancing the biological relevance and broader applicability of the findings. Importantly, LWR analyses of T. anak conducted across multiple locations and temporal scales, while explicitly incorporating environmental variability and aquaculture management practices, would yield more robust and transferable growth parameters. Further research incorporating seasonal variability, sex-specific analyses, and environmental parameters is recommended to better understand the factors influencing growth patterns and condition in this species. Such approaches would enhance the utility of LWR data to ecological research, stock assessment, and the sustainable development of pompano aquaculture.
Conclusion
Accurate species identification of T. anak has been hindered by mislabeling of reference sequences in global databases. In this study, mitochondrial COI sequences from morphologically identified T. anak collected in northern Vietnam were analyzed to resolve this taxonomic ambiguity and to confirm its phylogenetic placement among 16 other Trachinotus congeners. The generated sequences of T. anak exhibited substantial interspecific divergence (15.2%) difference from T. ovatus and recovered as a distinct lineage in the phylogenetic analyses. The species delimitation and haplotype network analyses further confirmed discrete MOTUs and clear species boundaries of both T. anak and T. ovatus. Additionally, analysis of growth patterns revealed a significant negative allometric trend, indicating that body weight increases at a proportionally lower rate than body length. The regression analysis demonstrated a strong correlation between length and weight, suggesting that body length serves as a reliable predictor of body mass for this species under the prevailing culture conditions. The observed deviation from isometric growth may be attributed to the combined effects of environmental factors, feeding strategies, stocking density, and physiological processes associated with mariculture practices. These findings provide essential baseline information for elucidating growth dynamics, evaluating body condition, and facilitating improved stock management and aquaculture optimization of T. anak. Overall, these findings resolve the long-standing species identification issue and provide a foundation for the sustainable monitoring and management of T. anak in mariculture systems in Vietnam.