
Introduction
Butyric acid (BA, C4H8O2) like short-chain fatty acids (SCFAs) that are produced through the metabolism of various substrates by anaerobic bacteria and are abundant in the gut of aquatic animals, serving as a primary energy source (Tran et al., 2020). Among SCFAs, BA, its salts, and their protected forms have been reported to possess growth-promoting, immune-stimulating, and anti-oxidative properties that are beneficial in the scope of nutrition of aquaculture sector (Abdel-Latif et al., 2020). The most abundant and stable forms of BA are its salts and esters (Jiang et al., 2018). Importantly, these compounds have shown positive effects when supplemented in animal feed (Abdel-Latif et al., 2020), with butyrate glycerides like mono-butyrin and tributyrin (TB, C15H26O6) those are recently gained the interest of researchers as potential supplements in fish and crustacean feeds (Palma et al., 2023).
TB, one of the stable forms of butyrate glycerides, is a non-volatile and nearly odorless compound with better palatability than other BA derivatives (Palma et al., 2023). Dietary TB has been reported to promote gut health, immune system, and the digestive enzyme activity in several fish and crustacean species. Improved growth performance and enhanced immunity have been reported in snakehead fish (Channa argus), black seabream (Acanthopagrus schlegelii), grass carp (Ctenopharyngodon idellus), blunt snout bream (Megalobrama amblycephala), and common carp (Cyprinus carpio) after the supplementation of TB in their diets (Hou et al., 2019; Hu et al., 2021; Liang et al., 2021; Volatiana et al., 2020; Xie et al., 2021). Studies by Hou et al. (2019) and Volatiana et al. (2020) noted an elevation in digestive enzyme activities in snakehead fish and black sea bream under the influence of dietary TB. Furthermore, blunt snout bream fed with TB-supplemented diets showed higher survival rates when challenged with Aeromonas hydrophila (Liang et al., 2021).
Among the crustacean species cultured worldwide, Pacific white shrimp (Penaeus vannamei) stands at the top in terms of production (FAO, 2022). Therefore, investigating the effects of multi-purpose feed additives such as TB in Pacific white shrimp culture has recently garnered increasing attention. Lee et al. (2021), Liu et al. (2021), Ding et al. (2022) and Rairat et al. (2023) have reported increases in growth and immunity-related parameters in groups of shrimp fed with TB-added diets in addition to stimulating the digestive enzyme activity (Ding et al., 2022; Liu et al., 2021), immune-related gene expression (Ding et al., 2022; Lee et al., 2021), improving intestinal morphology (Liu et al., 2021) and the apparent digestibility coefficient of lipid (Lee et al., 2021; Rairat et al., 2023) in the diet. Lee et al. (2021) conducted a challenge test against Vibrio harveyi for Pacific white shrimp fed with TB-supplemented diets, reporting a significantly higher survival rate at the 0.2% inclusion level. Those studies emphasize selected aspects such as growth and immunity or digestive physiology, and few have jointly evaluated nutrient digestibility, immune-related gene expression, and disease resistance outcomes within a single integrated study. However, previous literature has not suggested an optimum dietary TB inclusion level for crustacean species, despite having tested various levels. Additionally, less attention has been given to the use of TB as an effective therapeutic agent against bacterial diseases in shrimp culture. Recent studies have highlighted the severe losses in shrimp culture caused by acute hepatopancreatic necrosis disease (AHPND) in Asian countries, with Vibrio parahaemolyticus which was identified as the main causative agent (Kumar et al., 2020).
Therefore, a more comprehensive understanding of how dietary TB influences multiple physiological responses including growth, feed utilization, digestive enzyme activity, antioxidant and immune function, nutrient digestibility, and immune-related gene regulation under controlled experimental conditions is needed. Furthermore, determining an optimal dietary inclusion level will be valuable for practical feed formulation. The present study aimed to investigate the effects of graded levels of dietary TB on growth performance, feed utilization, innate immunity, digestive function, nutrient digestibility, immune- and growth-related gene expression, and disease resistance against V. parahaemolyticus in P. vannamei.
Materials and Methods
A basal diet (Con) was prepared (20% fish meal) according to the nutrient requirement of P. vannamei. TB (60% purity, Feedbest, Cheonan, Korea) was incorporated into the basal diet at product inclusion levels of 0.05%, 0.10%, and 0.20%, corresponding to 0.03%, 0.06%, and 0.12% active TB equivalent, respectively. These diets were designated as TB5, TB10, and TB20. Ingredients were mixed thoroughly while adding 10% distilled water and fish oil then, pelleted (size 2 mm) through a pelleting machine and pellets were dried with an electric dryer at 25°C for 8 hours. Table 1 shows the experimental diet formulation and their proximate compositions.
Mineral mix contains (g kg-1): MgSO4·7H2O, 80; NaH2PO4·2H2O, 370; KCl, 130; Ferric citrate, 40; ZnSO4∙7H2O, 20; Ca-lactate, 357; CuCl, 0.2; AlCl3∙6H2O, 0.15; Na2Se2O3, 0.01; MnSO4∙H2O, 2; CoCl2∙6H2O, 1.0.
Vitamin premix contains (g kg-1): L-ascorbic acid, 121; DL-α tocopheryl acetate, 19; thiamin hydrochloride, 2.7; riboflavin, 9.1; pyridoxine hydrochloride, 1.8; niacin, 36; Ca-D-pantothenate, 12.7; myo-inositol, 182; D-biotin, 0.27; folic acid, 0.68; p-aminobenzoic acid, 18; menadione, 1.8; retinyl acetate, 0.73; cholecalciferol, 0.003; cyanocobalamin, 0.003.
The shrimp for the experiment were obtained from a private hatchery in Shinan, Jeollanam-do, Korea and transferred to the Jeju National University, Marine Science Institute. They were acclimated in rearing tanks for two weeks and fed with a commercial diet (Woosung Feed No. 1–3, protein: 40%, lipid: 6%) during the period. For the feeding trial, juvenile shrimp (0.50 ± 0.0 g) were distributed randomly to 16 tanks (240 L) with 25 shrimp per tank. The complete randomized design was applied to assign diets for quadruplicate groups of treatments and shrimp feeding was done 6%–10% of the body weight for eight weeks at six times a day (08:00, 10:00, 12:00, 14:00, 16:00 and 18:00 h). The feeding rate was adjusted weekly based on bulk weighing of each tank. Feed intake was assumed to be fully consumed as no uneaten feed was detected during routine observations. Continuous aeration was provided to all rearing tanks. At every three days, water exchanging was done with preheated filtered seawater. Water quality parameters including temperature, dissolved oxygen, salinity, and pH, were measured daily, while total ammonia was monitored three times per week. No significant differences were detected among treatments and values were: temperature 30.0 ± 1.50°C, dissolved oxygen 6.10 ± 0.59 ppm, salinity 32.1 ± 0.32 ppt, pH 7.24 ± 0.17 and ammonia 0.04 ± 0.05 ppm.
At the end of the 8-week feeding trial, total count and the bulk weight of the experimental shrimp were measured in each tank. Each rearing tank was considered the experimental unit, and all statistical analyses were performed using tank-level mean values. Growth performance and feed utilization indices were calculated as follows:
After weighing, 12 shrimp from each dietary group (3 shrimp per tank) were randomly captured and anaesthetized with ice-cold water. After that, the hemolymph was collected using a 1 mL syringe containing Alsever’s solution at a ratio of 1:1, centrifuged (800×g, 20 minutes, 4°C), and separation of the supernatant was done and used for analysis. Hemolymph was stored in a freezer (–80°C) until analysis. The sampled shrimp were used for whole-body composition analysis.
Experimental diet and whole-body proximate compositions were analyzed in accordance with AOAC (2019) methods. Moisture was analyzed by oven-drying at 125°C for 3h and ash was analyzed by direct incineration method (550°C, 4 h). Crude protein was analyzed by automatic protein analyzer (Kjeltec 2300, FOSS, Hilleroed, Denmark) and lipid level was quantified by the chloroform-methanol extraction method as described by Folch et al. (1957). Aspartate aminotransferase (AST) and alanine aminotransferase (ALT) levels, total protein, and glucose in the hemolymph were measured using an automatic biochemical analyzer (SLIM, SEAC, Florence, Italy). For statistical analysis, values from three shrimp within the same tank were averaged, and each tank was treated as the experimental unit.
Hemolymph from three shrimp was sampled from each tank, and individual measurements were averaged to obtain a single tank-level value and each tank was considered the experimental unit for statistical analysis. Respiratory burst activity (oxidative radical production by phagocytes) in hemolymph (NBT, nitro-blue tetrazolium) was performed according to the method described by Zhang et al. (2013). A turbidimetric assay was done to analyze the lysozyme activity following Hultmark et al. (1980). Phenoloxidase (PO) activity was analyzed following the methods described by Hernández-López et al. (1996). Superoxide dismutase (SOD) and glutathione peroxidase (GPx) activities were determined by using commercial assay kits as SOD colorimetric assay Kit (DG-SOD400, Dogenbio, Seoul, Korea) and GPx colorimetric assay kit (703102, Cayman chemical company, Ann Arbor, MI, USA) respectively. Analysis of anti-proteases activity in hemolymph was done following the method described by Ellis (1990).
The hepatopancreas was selected as the target tissue for gene expression analysis because it serves as a central organ for nutrient metabolism, digestive enzyme secretion, detoxification, and plays a key role in systemic immune regulation in shrimp. Hepatopancreases were collected from three shrimp within each tank, and relative expression values were averaged at the tank level. The tank was considered the experimental unit for statistical analysis. Collected hepatopancreases were used to extract RNA using TRI-zol® reagent (Sigma-Aldrich, St. Louis, MO, USA). RNA concentration and purity were measured using a μDropTM Plate (Thermo Scientific, Waltham, MA, USA). Synthesizing of cDNA was done using the Prime ScriptTM (TaKaRa Bio, Shiga, Japan) first-strand cDNA synthesis kit by adjusting the separated RNA 2.5 μg and the reaction mixture to a total volume of ٢٠ μL. Quantitative polymerase chain reaction (qPCR) was performed for one cycle at 95°C for 10 s, followed by 45 cycles at 95°C for 5 s, 58°C for 20 s, and 60°C for 30 s, as mentioned in Lee et al. (2021) using a real time PCR system (TP 950 Thermal Cycler DiceTM, TaKaRa Bio).
Relative mRNA expression levels of prophenoloxidase (proPO), crustin, insulin-like growth factor binding protein (IGF-BP), inhibitor of nuclear factor kappa-β kinase subunit beta (IKBK), transforming growth factor beta (TGF-β) and toll-like receptor 3 (TLR3) were calculated using the comparative 2−ΔΔCt method (Livak & Schmittgen, 2001). Primer pairs were designed and optimized to ensure comparable amplification performance, and amplification efficiencies of target and reference genes were verified to be similar and close to 100%. β-Actin and elongation factor 1-α (EF1-α) were used as reference genes, and the geometric mean of their expression levels was applied as the normalization factor following the recommendations of Vandesompele et al. (2002). Amplification specificity was confirmed by the presence of a single peak in melting curve analysis and the absence of non-specific products. The expression stability of β-actin and elongation factor 1-α across experimental treatments was evaluated and confirmed to be suitable for normalization. The primer sequences of the related genes are shown in Table 2.
| Target gene | Primer sequences (5’ to 3’) | Accession number/reference |
|---|---|---|
| β-Actin |
(F): 5’-GAGCAACACGGAGTTCGTTGT-3’ (R): 5’-CATCACCAACTGGGACGACATGGA-3’ |
AF300705.2 |
| EF1-α |
(F): 5’-GAAGTAGCCGCCCTGGTTG-3’ (R): 5’-CGGTTAGCCTTGGGGTTGAG-3’ |
XM027373349.1 |
| proPO |
(F): 5’-TTCAACGGTAGACCCGTGATTCTTC-3’ (R): 5’-TCTTGCCGGGTTTAAGGTGAACAGT-3’ |
AY723296.1 |
| Crustin |
(F): 5’-CTTGCACACGTGTTCTCCCAAACA-3’ (R): 5’-ACCAAGATACTCGACTGCCCACAA-3’ |
AY486426.1 |
| IGF-BP |
(F): 5’-GTGGGCAGGGACCAAATC-3’ (R): 5’-TCAGTTACCACCAGCGATT-3’ |
Lee et al. (2021) |
| IKBK |
(F): 5’-GCAAGAACCGCACAACACAT-3’ (R): 5’-AAGCATTAGGAACGGGCTGG-3’ |
JN180642.1 |
| TGF-β |
(F): 5’-CGCATACACCTATACGAGAG-3’ (R): 5’-CTGGTGCTGAGTCATATCTG-3’ |
MH259763.1 |
| TLR3 |
(F): 5’-TGACGACCTGCATGAACACCTCTT-3’ (R): 5’-GGTGCCACGGTACACGAAACATAAC-3’ |
JN180638.1 |
Diets for the digestibility trial were prepared by adding 1% chromium oxide (Cr2O3) as an inert marker to the experimental diets. The digestibility trial was conducted as a separate experiment using another group of shrimp (mean initial body weight: 9.50 ± 0.5 g). Shrimp were randomly distributed into triplicate tanks per dietary treatment (25 shrimp per tank), and each tank was considered the experimental unit for statistical analysis. Feces collection was done daily through siphoning two times and separation of feces were done by filtering with a filter paper and kept at –20°C until freeze-dried. Proximate composition analysis of feed and feces was performed according to the AOAC (2019) methods. The chromium oxide contents of digestibility diets and shrimp feces were analyzed following Divakaran et al. (2002). The equations used to calculate apparent digestibility coefficients (ADCs) of experimental diets are as follows:
After the feeding trial, the hepatopancreas from 12 shrimp per experimental group (3 shrimp per tank) were extracted and analyzed for digestive enzyme activity. Values within the same tank were pooled and averaged, and each tank was treated as the experimental unit for statistical analysis. Trypsin activity was assayed using benzoyl-DL-arginine-p-nitroanilide (BAPNA) as a substrate and chymotrypsin activity was analyzed using Suc-Ala-Ala-Pro-Phe-p-nitroanilide (SAPNA) as the substrate according to Erlanger et al. (1961). Amylase activity was measured according to the method described by Worthington (1993) using 1% starch solution as the substrate. Analyzing lipase activity was done following the spectrophotometric method described by Kuepethkaew et al. (2017) using p-nitrophenyl palmitate (p-NPP) as a substrate.
In order to investigate the bacterial disease resistance of P. vannamei fed with experimental diets, a challenge test was done using V. parahaemolyticus strains. The V. parahaemolyticus strain used in this study was 13-028/A3, obtained from the Korean Culture Collection of Aquatic Microorganisms (Busan, Korea). The isolate originated from AHPND-affected shrimp in Vietnam, and its pathogenicity was verified by duplex PCR targeting pirA and pirB virulence genes. The challenge test was performed in an isolated quarantine setup at the Marine Research Institute of Jeju National University. At the end of the feeding period, shrimp from the four replicate feeding tanks within each dietary treatment were pooled by diet to eliminate potential carry-over tank effects from the growth phase. Shrimp were then randomly redistributed into three independent challenge tanks per dietary treatment (120 L acrylic tanks; 13 shrimps per tank). Each challenge tank was considered the experimental unit for survival analysis. Tryptic soy broth (211825, BD DifcoTM, Franklin Lakes, NJ, USA) was used to cultivate the bacteria with 2% NaCl. Incubation was done while shaking at 150 rpm for 24 h at 30°C to reach 1 × 107 CFU/mL. After inoculating each tank with 40 mL of bacterial culture solution, mortality was observed for 170 hours. Water temperature in the tanks was kept at 30°C during the challenge test period and an air generator was installed to maintain continuous aeration. To maintain water quality while minimizing uncontrolled stress, water exchange (80%) was performed once every 24 hours after infection establishment and shrimp mortalities were recorded every hour.
Completely randomized design was used to assign the dietary treatments. Data were subjected to one-way ANOVA. When ANOVA identified the differences among groups, the mean differences were compared using Tukey’s honestly significant difference (HSD) multiple range test. Statistical significance was determined at p < 0.05. A follow-up trend analysis by orthogonal polynomial contrasts was run to determine whether the effects are linear and/or quadratic. Data was presented as mean ± SD and percentage data were arcsine transformed before the statistical analysis. Survival following V. parahaemolyticus challenge was analyzed using Kaplan-Meier survival curves, with differences among dietary treatments assessed by the log-rank test. In addition, Cox proportional hazards regression was performed to estimate hazard ratios for mortality among dietary treatments relative to the control diet. All the analyses were conducted using SPSS version 24.0 (IBM, Armonk, NY, USA).
Results
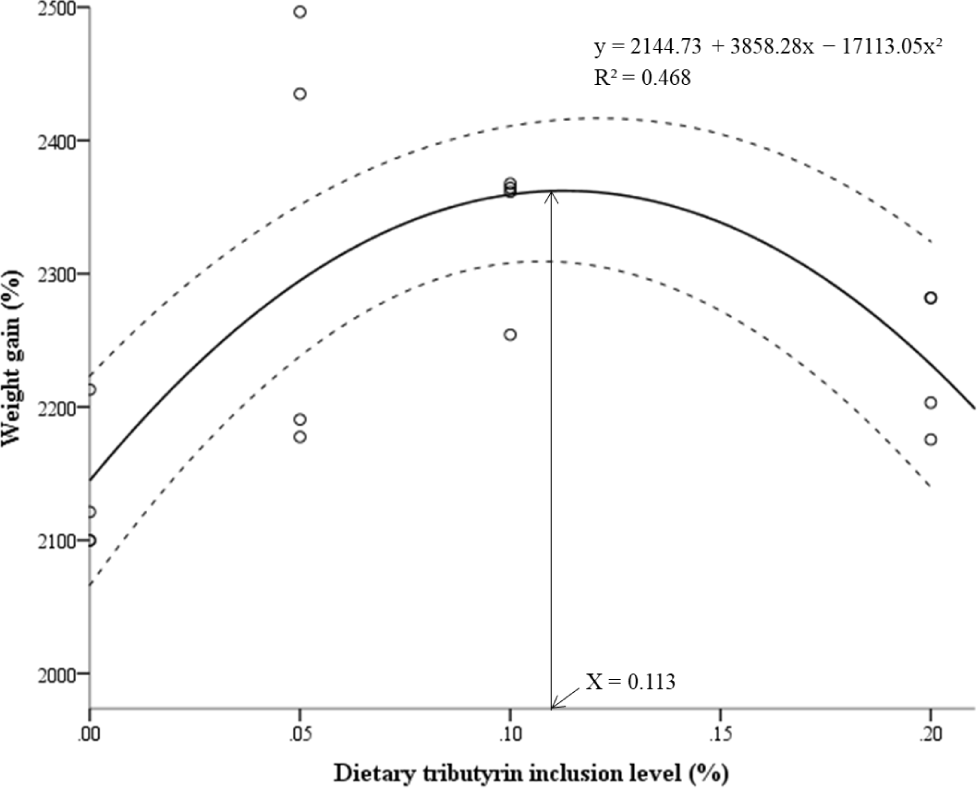
The weight gain of the TB10 group was significantly higher than that of the Con group (Table 3). The SGR was significantly higher in the TB10 group than that in the Con group (p < 0.05). FCR was significantly lower in TB5 and TB20 compared to the Con group (p < 0.05). PER was significantly higher in the TB5 group compared to the Con group (p < 0.05). Survival rates of all experimental groups were standing over 91%, and no significant difference was observed between the Con group and the TB added groups (p > 0.05). Whole-body compositions of protein, dry matter, ash and lipid contents were not significantly affected (p > 0.05) by dietary TB supplementation (Table 4). Polynomial regression indicated a significant quadratic effect of dietary TB level on weight gain of P. vannamei (F = 5.720, df = 2,13, p = 0.017, R² = 0.468). The maximum weight gain was recorded at 0.113% (95% CI: 0.00%–0.19%) of TB level (Fig. 1).
Hematological parameters analysis showed no significant difference (p > 0.05) in ALT, AST, glucose, and total protein levels between the Con group and the TB-added group (Table 5). As a result of the non-specific immunoassay (Table 6), NBT levels were significantly higher in all of the TB-added groups compared to the Con group (p < 0.05). PO levels and lysozyme activities were significantly higher in TB10 and TB20 than in the Con group (p < 0.05). Anti-protease levels were significantly higher in TB10 and TB20 than in the Con group (p < 0.05). As a result of antioxidant enzyme activity analysis (Table 6), GPx activity was significantly higher in the TB10 and TB20 groups than that of the Con group (p < 0.05). SOD activity levels were significantly higher in all TB-added groups compared to the Con group (p < 0.05).
ADC of the diets were affected significantly by dietary TB supplementation (Table 7). Dry matter digestibility and protein digestibility were significantly elevated in the TB-added group than those of the Con group, and significantly higher in the TB20 group than in the TB5 and TB10 groups (p < 0.05). Lipid digestibility was significantly higher in the TB20 group than that of the Con group (p < 0.05).
According to the results of digestive enzyme activity analysis, trypsin activity was significantly higher in TB10 and TB20 groups compared to the Con group (p < 0.05) (Table 8). In comparison of Con and TB-added groups, there was no significant difference observed in chymotrypsin activity (p > 0.05). Activity of amylase was significantly higher in the TB20 group than in the Con group (p < 0.05).
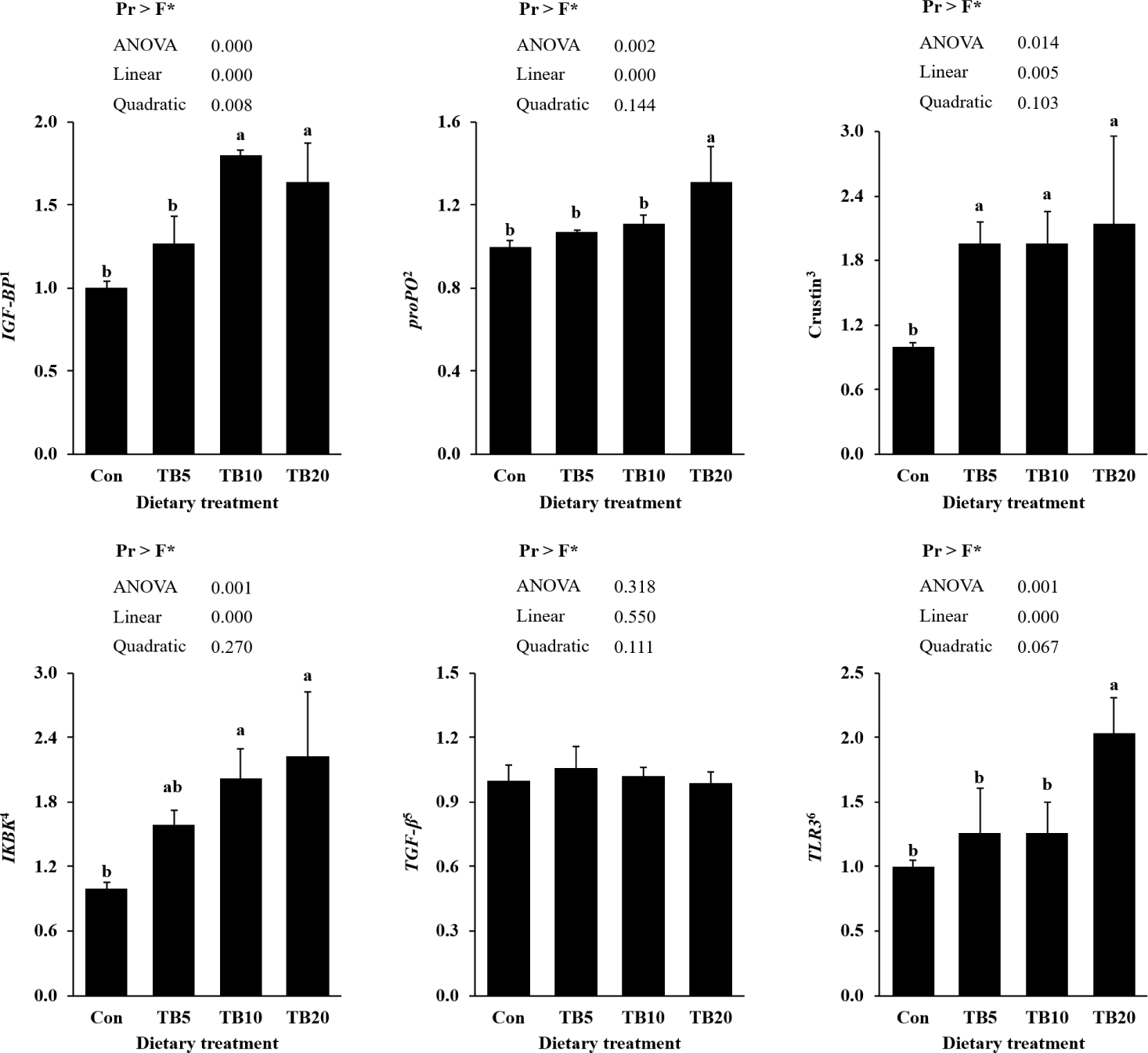
Relative expression of growth, inflammatory and immune genes were affected significantly (p < 0.05) by dietary TB supplementation (Fig. 2). The expression level of IGF-BP was significantly higher in the TB10 and TB20 groups than in the Con group (p < 0.05). Expression level of proPO was elevated significantly in the TB20 group than the Con, TB5, and TB10 groups (p < 0.05). Crustin expression levels were significantly higher in the TB-added group than those of the Con group (p < 0.05). The expression level of IKBK was significantly higher in the TB10 and TB20 groups than that of the Con group (p < 0.05). TLR3 gene expression levels were significantly higher in all TB-supplemented groups compared to the Con group (p < 0.05), where the expression levels of TGF-β were not significantly different among all the treatments (p > 0.05).
According to the results of quadratic regression analysis using TB content in feed and WG%, the estimated optimum TB content was 0.113% in shrimp feed (Fig. 2).
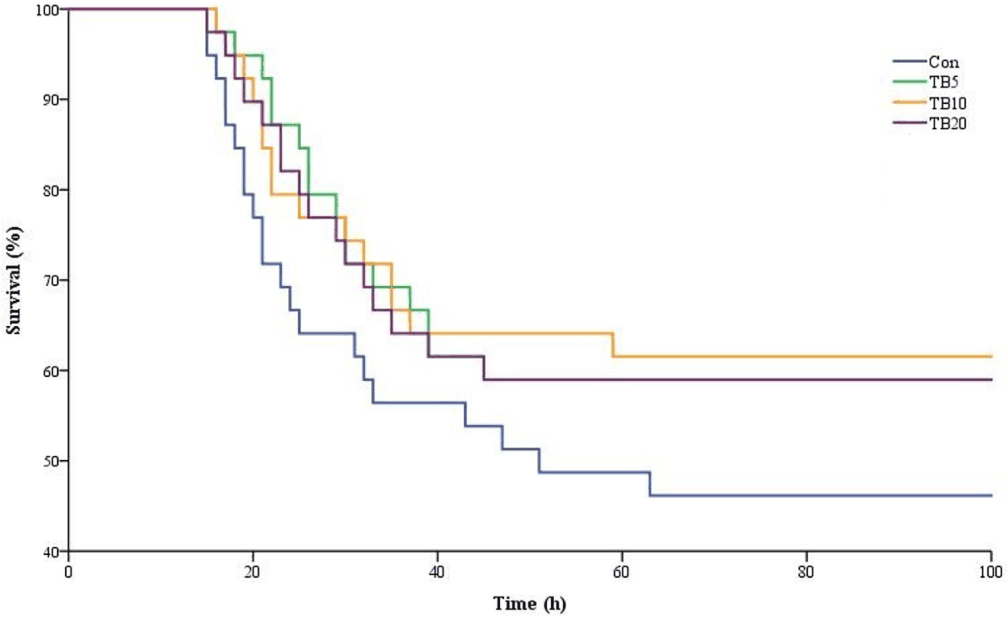
In the challenge test, deaths were observed from 15 hours after the immersion of the bacterial suspension, and the last death was observed at 63 hours of the test (Fig. 3). Kaplan-Meier survival analysis showed no significant differences in survival probability among dietary treatments during the challenge period (log-rank test: χ² = 2.97, df = 3, p = 0.397). Cox proportional hazards regression revealed no significant effect of dietary treatment on mortality risk (Table 9). Compared with the control, hazard ratios were 1.50 for TB5, 0.97 for TB10, and 0.93 for TB20, with all 95% confidence intervals overlapping unity (p > 0.05).
Discussion
Within the gut of animals, butyrate compounds can be naturally produced by bacteria through the fermentation of ingested carbohydrates (Palma et al., 2023). These compounds are among the most important metabolites for epithelial cells in the colon (Bedford & Gong, 2018). Butyrate glycerides such as TB are composed of numerous BA molecules linked to a glycerol backbone, which are released in the gut and serve as a substrate for lipase (Palma et al., 2023). Earlier studies have reported that the availability of butyrate in the gut may enhance the growth performance of fish and shrimp through indirect effects, such as the development of the intestinal tract, modulation of the gut microbiota, and improved antioxidant activity (Abdel-Latif et al., 2020). In line with the TB10 diet group in the current study, Ding et al. (2022) and Rairat et al. (2023) reported a significant improvement in final body weight (FBW) and WG with 0.1% TB (purity 98%) supplementation in shrimp diets. Similarly, Lee et al. (2021) observed significant enhancements in WG and SGR with 0.2% TB (purity 60%) inclusion. Considering the growth performance results, dietary TB (60% purity) supplementation showed a positive trend across all supplemented groups, with a significant improvement observed specifically in the TB10 group compared with the control, indicating that TB can promote shrimp growth at appropriate inclusion levels. Furthermore, the up-regulated expression of the IGF-BP gene, which plays a crucial role in regulating growth and metabolism (LeRoith et al., 1992), can serve as evidence for the growth-promoting effect of TB in Pacific white shrimp. Similarly, Lee et al. (2021) also observed a significant elevation in IGF-BP gene expression when TB was added to shrimp diets.
In the present study, the body composition of the shrimp was not affected by the TB supplementation, although Ding et al. (2022) reported higher crude protein and lipid content in TB-added groups. However, the authors pointed out that the observed increases in whole-body protein might be due to decreased protease activity in the intestine, leading to the deposition of more protein in the shrimp’s body. In our case, trypsin activities in the TB groups were higher, and chymotrypsin activities were comparable to those of the Con group. Therefore, no evident changes in whole-body composition were observed.
The activity of antioxidative enzymes in hemolymph such as SOD and GPx (Martínez-Álvarez et al., 2005) and activities of PO, NBT, Lysozyme (Subramanian et al., 2014) and antiprotease (Abarike et al., 2019) are considered as indicators of the innate immune system. In reference to the present study, dietary TB supplementation appears to have had a positive effect on shrimp innate immunity. Although the mechanisms through which TB modulates the crustacean immune system remain poorly understood, previous studies have suggested that SCFAs could modulate immune system function. In animals, the regulation of the immune system and metabolic disorders through the inhibition of histone deacetylases and the activation of G protein-coupled receptors has been reported to be influenced by SCFAs (Tran et al., 2020). SCFAs, including butyrate, undergo rapid absorption in the gut lumen and perform crucial immune-related functions such as participating in the inflammation process and systemic autoimmune responses (Sun et al., 2017). The increases in immune parameters observed in the 0.1% and 0.2% TB groups suggest a state of immune priming rather than basal activation, indicating that the shrimp were better prepared to respond to pathogenic challenges without experiencing chronic inflammation. Butyrate is known to modulate immune function by influencing signaling pathways and promoting reactive oxygen species-mediated antimicrobial defense. However, excessive activation could theoretically increase oxidative stress or trigger inflammatory responses. In this study, no such adverse effects were detected at the tested supplementation levels and collectively, these findings indicate that TB supplementation at 0.1%–0.2% enhances innate immune readiness while remaining within a physiologically beneficial range. Consistent with our findings, Lee et al. (2021), Liu et al. (2021), and Ding et al. (2022) reported that TB-supplemented diets significantly improved innate immunity in Pacific white shrimp.
Furthermore, confirming the findings on immune aspects of hemolymph, immune and inflammatory responses-related gene expression were upregulated in shrimp had TB-included diets. In particular, proPO expression was upregulated in TB-fed groups, consistent with the role of the proPO system as a key melanization-based defense mechanism against invading pathogens (Cerenius & Söderhäll, 2004). Expression of IKBK, a regulator of NF-κB activation in pro-inflammatory signaling (Karin & Delhase, 2000), and crustin, an antimicrobial peptide induced through Toll/Immune deficiency (IMD) pathways in response to microbial stimulation (Chen et al., 2019), also increased compared to the control diet, indicating that dietary TB may enhance the baseline immune preparedness of shrimp. Although TLR3 is primarily characterized for antiviral recognition, previous studies in P. vannamei have reported altered TLR expression during immune stimulation and bacterial challenge (Ding et al., 2022; Habib et al., 2021), suggesting its involvement in broader innate immune regulation. Therefore, in this study, TLR3 was selected as a representative TLR-related marker to evaluate whether TB supplementation modulates upstream immune signaling capacity rather than as a direct Vibrio-specific pattern-recognition receptor. The observed increase in TLR3 transcription implies a potential immunomodulatory effect of TB at the receptor-signaling level. Comparable with present study, elevated expression level of proPO was observed by Lee et al. (2021) and, upregulated Crustin and Toll genes were reported by Ding et al. (2022) in Pacific white shrimp which fed diets included TB. However, further validation using receptor-specific assays or downstream pathway tracking would be required to determine the mechanistic relevance of these responses.
The digestive capabilities of shrimp largely depend on the types, characteristics, and modulation of digestive enzymes, thus highlighting the importance of carefully evaluating the ingredients that are incorporated into formulated diets (Carrillo-Farnés et al., 2007). In our study, the ADCs of the diets (dry matter, protein and lipid) were significantly improved with 0.2% TB inclusion. Interestingly, hepatopancreas trypsin, amylase, and lipase activities followed the same pattern. TB is known to release butyrate through lipase activity, providing energy to gut epithelial cells and supporting digestion and nutrient absorption (Palma et al., 2023). Overall, TB appears to enhance diet digestibility due to its ability to upregulate digestive enzyme activity and improve intestinal health, which may also indirectly explain the enhanced growth performance in the TB groups in the present study. However, digestive enzyme activities and apparent digestibility coefficients continue to rise at the 0.2% TB supplementation level, where the quadratic regression results in weight gain, indicating that maximum growth is obtained at a 0.113% TB level. This discrepancy may reflect diminishing physiological returns, where further enhancements in digestive efficiency do not proportionally translate into somatic growth. One possible explanation is that the additional metabolic energy at higher TB levels may be redirected toward immune modulation or maintenance processes rather than growth. Because growth performance is influenced by multiple interacting factors, including nutrient digestibility, immune activity, antioxidant balance, and tissue deposition efficiency, the observed pattern suggests that the optimal supplementation level results from complex interactions among these physiological processes.
Several Vibrio species have been identified as causative agents in economically important bacterial diseases such as vibriosis and AHPND in shrimp aquaculture. Among these disorders, AHPND has recently emerged as the most concerning bacterial disease in shrimp, leading to substantial losses in shrimp farms, with V. parahaemolyticus having been identified as the main causative agent of AHPND (Kumar et al., 2020). Lee et al. (2021) investigated the protective effects of 0.1% and 0.2% TB-supplemented diets against V. harveyi. However, up to date, no previous studies have conducted V. parahaemolyticus challenge tests on shrimp fed with TB-supplemented diets, to the best of our knowledge. Consistent with the findings of Lee et al. (2021) the results of our challenge experiments clearly demonstrated the effective impact of TB on the survival of shrimp against V. parahaemolyticus. Organic acids, such as BA and its derivatives, are reported to have antimicrobial and bactericidal properties against certain infectious bacterial strains in aquatic species. Particularly, BA at lower concentrations has a higher inhibitory capacity compared to other organic acids (Nuez-Ortin et al., 2012). da Silva et al. (2013) characterized several Vibrio species (Vibrio alginolyticus, V. harveyi, Vibrio anguillarum and Vibrio campbellii) in the Pacific white shrimp gut and found that dietary sodium butyrate inhibited the growth of these pathogens without significantly suppressing the relative abundance of favorable lactic acid bacteria in the intestine. Similarly, BA-containing diets inhibited the growth of V. campbellii in Pacific white shrimp (Defoirdt et al., 2006). Moreover, Rairat et al. (2023) reported a significant reduction of Vibrio spp. count in the shrimp hepatopancreas and intestine after feeding TB-added diets.
The influence of dietary TB on the shrimp’s immune system may have also provided a protective effect against V. parahaemolyticus in the present study. Particularly, significantly higher NBT, lysozyme, SOD, and GPx activities were observed at the 0.1% TB inclusion level, which exhibited the highest cumulative survival rate in the challenge experiment. However, due to the scarcity of related literature, additional investigations are required to gain further insights into the mechanisms through which TB modulates the disease resistance and immune response in shrimp at the cellular level. Given the outstandingly positive impacts of TB inclusion in Pacific white shrimp diets, an optimum level for dietary inclusion is worth recommending. However, previous limited studies have not determined an exact optimum level and have instead recommended a range of levels. Based on our calculations using a quadratic regression model, highest weight gain can be achieved at a 0.11% TB inclusion level, which is slightly lower compared to the ranges recommended in previous studies. However, it should be noted that this optimum reflects growth performance specifically where some physiological indices, including digestive enzymes and immune-related parameters, continued to show incremental improvements at 0.2% TB, suggesting a dose-responsive enhancement in certain functional traits. Nevertheless, when the overall dataset is considered, the 0.10%–0.11% inclusion range appears to provide a balanced and practical supplementation rate. This indicates that moderate TB levels are sufficient to achieve both performance and health benefits without requiring the highest tested inclusion level.
Conclusion
Collectively, our findings confirmed that dietary TB supplementation improved growth performance and feed utilization, enhanced immune- and growth-related gene expression, promoted innate immune responses, increased nutrient digestibility, and stimulated digestive enzyme activity in Pacific white shrimp. Furthermore, TB supplementation improved disease resistance against V. parahaemolyticus even at the 0.10% inclusion level. Based on quadratic regression analysis, an estimated optimal dietary TB level of approximately 0.113% is recommended to achieve maximal growth performance in P. vannamei under the present conditions.