





Background
The order Bangiales (Nägeli 1874) is an order of distinctive and morphologically simple red algae that represents an ancient lineage (Butterfield 2000). The foliose Bangiales includes the most highly valued seaweeds grown in aquaculture from Korea, Japan, China, and Southeast Asia for the past several hundred years (Mumford and Miura 1988). This order has been considered monophyletic with the single family Bangiaceae. Traditionally, Bangia Lyngbye and Porphyra C. Agardh have been recognized on the basis of gametophyte morphology: unbranched uni- to multiseriate filaments in the genus Bangia and blade in the genus Porphyra (Oliveira et al. 1995). Recently, the revision of the order Bangiales, based on the phylogeny using nrSSU and rbcL genes, has greatly improved the taxonomic understanding of this group (Sutherland et al. 2011). According to Sutherland et al. (2011), the order Bangiales was split into eight of foliose (Boreophyllum, Clymene, Lysithea, Miuraea, Porphyra, Pyropia, and Wildemania) and seven of filamentous genera.
The genus Miuraea was established by Sutherland et al. (2011) based on molecular analyses and composed of only one species, M. migitae (N.Kikuchi, S.Arai, G.Yoshida & J.A.Shin) N.Kikuchi, S.Arai, G.Yoshida, J.A.Shin & M.Miyata. This species was originally described as Porphyra migitae, based on the specimen collected from Misaki town in Osaka Prefecture, Japan (Kikuchi et al. 2010). M. migitae was distinguished from other foliose Bangiales species by growing on the subtidal habitat and having fire red to pink color (Kikuchi et al. 2010). The distribution of M. migitae was reported only from Japan (Osaka and Nagasaki), attached to the dead bivalve shell and rope (Sutherland et al. 2011).
In Korea, the study of Porphyra has been carried out on morphology, physiology, flora, and culture by several authors (Hwang and Lee 2001; Kim and Kim 2011). To date, seven species of Porphyra and 13 species of Pyropia J.Agardh (previously known Porphyra from Korea) have been reported from Korea, mostly collected in the intertidal zone (Kim et al. 2013). Two species, Pyropia koreana (M.S. Hwang & I.K. Lee) M.S. Hwang, H.G. Choi, Y.S. Oh & I.K. Lee and Pyropia yezoensis (Ueda) M.S. Hwang & H.G. Choi, are known from subtidal habitats (Hwang and Lee 2001; Kim and Kim 2011). However, the specimens of the order Bangiales from subtidal habitats have been scarcely explored because of the difficulty of sampling in this environment, despite the possibility of discovering new or unrecorded species.
In this study, we collected foliose Bangiales specimens from a depth of 15 m in Udo, Jeju Island, Korea. We conducted morphological observations and rbcL sequence analysis in order to confirm the taxonomic position of these subtidal specimens. This is the first record of the occurrence of the order Bangiales species from under 15 m depth of subtidal habitats in Korean coasts.
Methods
Four samples were collected by SCUBA diving from a depth of 15 m of Udo (33° 30′ N, 126° 55′ E), Jeju Island, Korea, in 9 May 2015 (E15019) and 17 May 2016 (E16064, E16065, E16066). Thalli were mounted on herbarium sheets as voucher specimens. Some parts of thallus were cutoff to stain with 1% aqueous aniline blue acidified with a drop of 1% HCL and to make permanent slide in a solution of 50% Karo syrup for microscopic observations. Photomicrographs were taken with a Canon EOS 600D digital camera mounted on the Olympus BX43 microscope. Voucher specimens were deposited in the herbarium of Jeju National University, Korea (JNUB).
Genomic DNA was extracted from the specimens using the DNeasy Plant Mini Kit (Qiagen, Hilden, Germany) following the manufacturer’s instruction. The primer pairs for rbcL gene were rbcLJNF1-rbcLJNR1 and rbcLJNF2-rbcLJNR2 (Kang and Kim 2013). Amplification condition for rbcL consisted of 7 min at 97 °C for pre-denaturation, followed by 45 cycles of 1 min at 97 °C, 1 min 47 °C, and 2 min at 72 °C, with a final 7 min extension cycle at 72 °C, and a soak cycle at 4 °C. PCR products were purified using the AccuPrep® PCR Purification Kit (Bioneer, Daejeon, Korea) following the manufacturer’s instructions. Nucleotide sequences of rbcL were determined on strands of PCR amplification products at the Macrogen sequencing facility (Macrogen Inc., Seoul, Korea). The sequences were edited using Mega ver. 7.0.14 (Tamura et al. 2013), and multiple sequence alignments were constructed with 39 taxa of the order Bangiales from GenBank with two taxa of Hildenbrandiales as out-group.
Maximum likelihood (ML) analysis was conducted using RAxML software (Stamatakis 2006) with the GTR + I + Γ evolutionary model to confirm the taxonomic position of Korean specimens. We used 1000 independent tree inferences using the “-#” option to generate bootstrap values for phylogenetic analysis with the algorithm “a” of “-f” option to search the best-scoring tree in one program run. The other options were used as a default set of the software.
Results
In total, 1174 base pairs of rbcL were aligned with 444 variable positions (37.8%) and 353 phylogenetically informative positions (30.0%). The Korean specimens (GenBank accession number KX587462-KX587465) were almost identical with M. migitae from Japan (EU521643), showed 0.2% sequence divergence (Fig. 1). In addition, M. migitae was clearly separated from other Korean specimens, Pyropia koreana (HQ728198), Py. kuniedae (HQ728200), Py. pseudolinearis (HQ718196), and Py. yezoensis (AB366140 and GQ427218), by 10.2–10.5%, 10.1–10.3%, 9.4–9.7% and 11–11.1% sequence divergences, respectively.
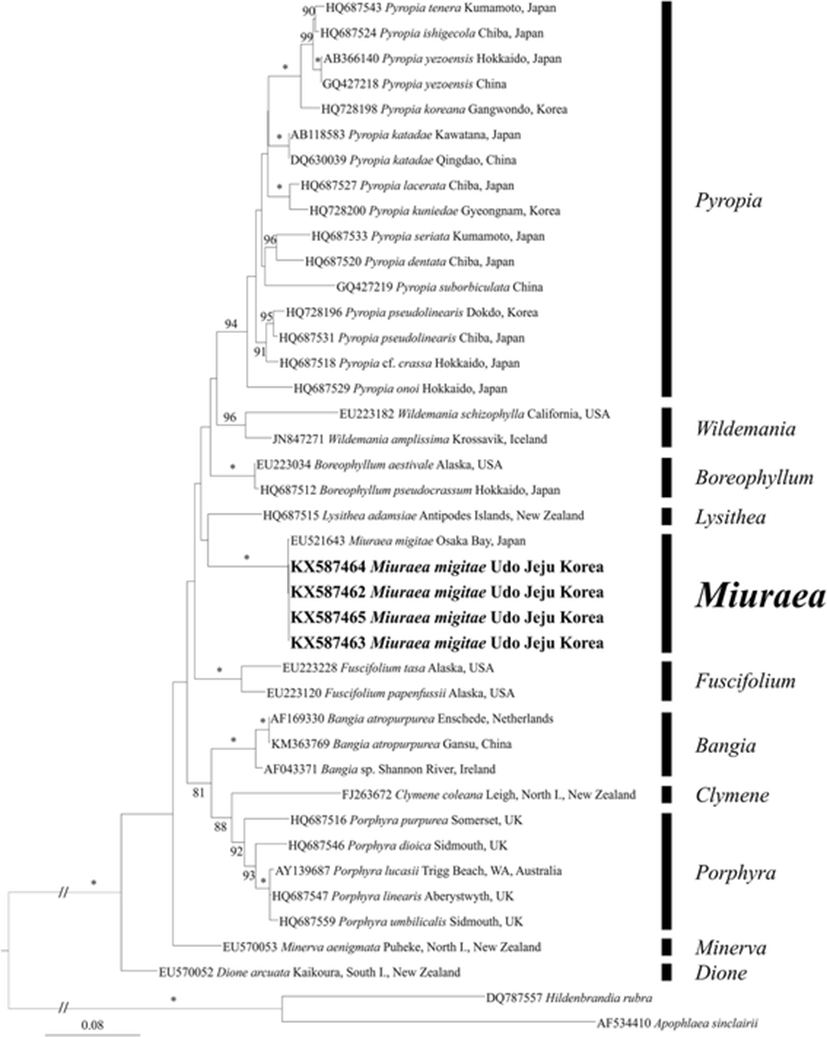
The clade of the genus Miuraea formed a monophyletic group with strong support, whereas there was a sister genus Lysithea without support. The monophyly of each genus was supported by strong bootstrap values (100% for Bangia, Boreophyllum, Fuscifolium, and Miuraea; 96% for Wildemania; 94% for Pyropia; and 92% for Porphyra). Pyropia formed a clade with Wildemania and Boreophyllum, but not supported. Porphyra was related to Clymene (a bladed species) and Bangia with filamentous morphology by moderate support values.
Miuraea migitae(N.Kikuchi, S.Arai, G.Yoshida & J.A.Shin) N.Kikuchi, S.Arai, G.Yoshida, J.A.Shin & M.Miyata
Basionym:Porphyra migitae N.Kikuchi, S.Arai, G.Yoshida & J.A. Shin
Type locality: Misaki town, Osaka Prefecture, Japan.
Holotype: SAP105477 (Kikuchi et al. 2010).
Korean name: Chambunhonggim (참분홍김).
Thalli are membranaceous, monostromatic, elliptic, obovate, and circular shape with cordate and rotund base, fire red to pink in color, and undulated entire margin (Fig. 2a, b). Thalli are up to 13 cm long, 6 cm wide, and 25–40 μm thick in the central portion (Fig. 2a, e). Vegetative cells are oblong, triangular, polygonal with rounded angles in the surface view, and 10–17 μm long × 7–10 μm broad in size (Fig. 2b, c). In the sectional view, cells are oblong and 15–20 μm high × 7–15 μm broad in size (Fig. 2e). Basal cells are 15–35 μm long × 6–20 μm broad in the surface view (Fig. 2d) and having rhizoidal filaments in both sides of thallus in the sectional view (Fig. 2f).
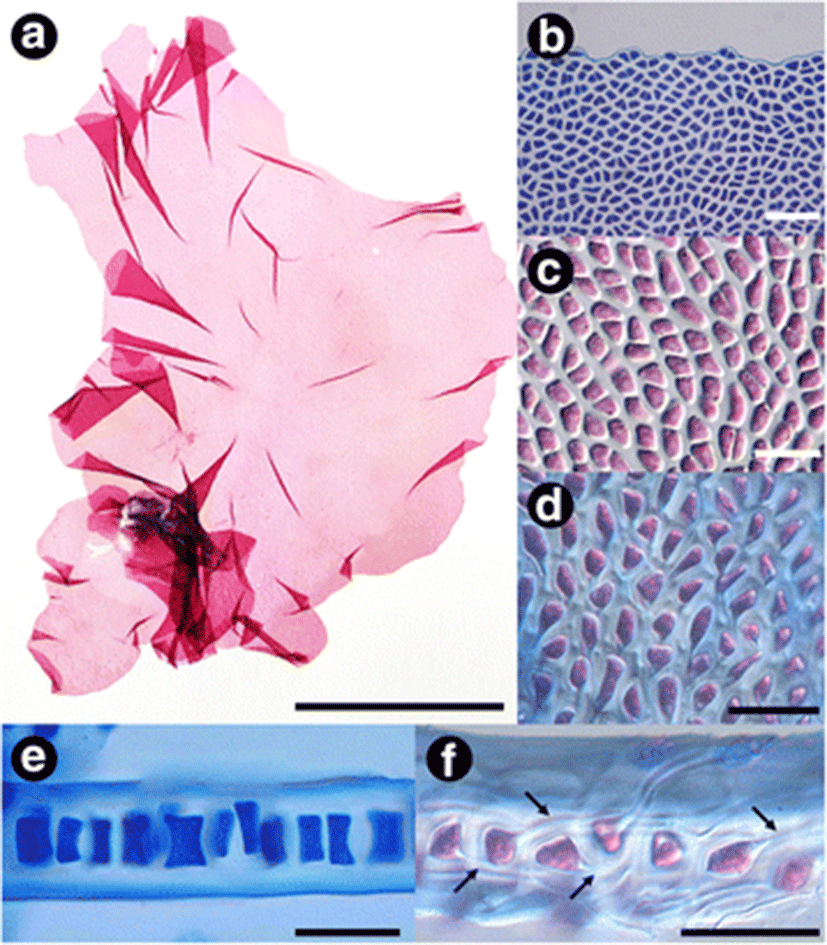
To date, M. migitae has been only collected from the subtidal zone in Udo, Jeju Island, Korea. The habitat is an area in a strong current and is covered with sand and a rhodolith bed.
Discussion
Miuraea migitae has been known as an endemic species in the subtidal zone of Japan. It is characterized by monostromatic thallus elliptic and obovate in shape, with fire red to pink color and 25 cm length × 13 cm width in size (Kikuchi et al. 2010). According to the culture study by Kikuchi et al. (2010), M. migitae can be distinguished from other subtidal species of Bangiales in Japan by the presence of asexual reproductive subcycles involving archeospores and neutral spores on the foliose thallus. Although our specimens were not observed any archeospores and neutral spores on thallus, Korean specimens were consistent with habitat, color, and vegetative characteristics in the description of M. migitae.
In recognizing the species of Pyropia, the vegetative features are viewed as important diagnostic characters, such as color, length, thickness, and projection of rhizoidal filaments including habitat (Table 1). M. migitae was found in the subtidal habitat like Py. koreana and Py. yezoensis with monostromatic thallus, but they can be distinguished from M. migitae by color, basal cell size, and the projection of rhizoidal filaments (both sides in M. migitae and one side in Py. koreana) (Table 1). Porphyra oligospermatangia, Pyropia katadae, and Py. tenera are having elliptic to obovate thallus with cordate base (Kim and Kim 2011), but they are different from M. migitae in length (24–47 cm in M. migitae and 30–56 cm in P. oligospermatangia), the thickness of thallus (25–40 μm in M. migitae, 40–50 μm in Py. katadae and 35–60 μm in Py. yezoensis), the habitats (subtidal zone in M. migiate and intertidal zone in P. oligospermatangia, Py. katadae, and Py. tenera), and the color (fire red to pink in M. migitae, yellowish to greenish in P. oligospermatangia, Py. tenera, and Py. yezoensis, and purple in Py. katadae) (Hwang and Lee 1994; Kim and Kim 2011).
Miuraea migitae | Porphyra oligospermatangia | Pyropia kuniedae | Py. koreana | Py. tenera | Py. yezoensis | |
|---|---|---|---|---|---|---|
Habitat | 6–15-m depth of subtidal zone | Intertidal zone | Low intertidal zone | Low intertidal to 5-m depth of subtidal zone | Upper to middle intertidal zone | Middle intertidal to 5-m depth of subtidal zone |
Color | Fire red, pink | Yellowish brown, pale-green toward the base | Purple, greenish red, brownish red | Bright red, brownish red | Yellowish red, brownish red, greenish red | Brownish red, greenish red |
Shape | Narrow elliptic, elliptic, obovate, circular | Elliptical, linear, lanceolate | Round, ovate | Elliptical, obovate | Elliptical, round, ovate, obovate | Round, ovate, obovate, oblanceolate, reniform |
Basal shape | Cordate, rotund, navel | Round, cordate | Cordate | Cuneate, round | Cuneate, round, cordate | Cuneate, round, cordate, umbilicate |
Dentation | Absent | Absent | Absent | Absent | Absent | Absent |
Undulation | Present | Present | Present | Present | Present | Present |
Length (cm) | 13–25 | 20–40 (−55) | 7–9.5 | 6–10 | 4.5–20 (−40) | 3.5–13 |
Width (cm) | 6–13 | 5–15 | 6.5–9 | 4–8 | 1.5–9 | 1–9 |
Thickness (μm) | 24–47 | 30–56 | 40–50 | 20–27 | 20–40 | 35–60 |
Vegetative cell shape | Oblong, triangular, polygonal | Circle, rectangle | Circle, rectangle, triangle | Rectangle | Rectangle | Rectangle |
Vegetative cell size (μm) | 8–17 × 6–10 | 7–11 × 6–9 | 12–15 × 7–10 | 13–17 × 9–10 | 15–21 × 9–12 | 13–19 × 7–14 |
Basal cell shape | Capitate | Elliptic | Elliptic, capitate | Elliptical | Circle, elliptic | Circle, elliptical |
Basal cell size (μm) | 15–35 × 6–20 | 18–42 × 13–21 | 22–27 × 13–16 | 22–25 × 9–12 | 22–33 × 13–21 | 22–33 × 10–23 |
Projection of rhizoidal filaments | Both sides | Both sides | Both sides | One side | Both sides | Both sides |
Division formula of spermatangia | 4 × 4 × 8 | 4 × 2 × 8 | 4 × 2–4 × 4–8 | 4 × 2–4 × 4 | 4 × 2–4 × 4–8 | 4 × 2–4 × 8–16 |
Division formula of zygotosporangia | 2 × 2 × 4 | 2 × 2 × 2–4 | 2 × 2 × 4 | 2 × 1 × 2 | 2–4 × 1–2 × 2–4 | 2 × 2 × 4 |
Reference | This study, Kikuchi et al. 2010 | Kim and Kim 2011 |
The molecular analyses demonstrated that the rbcL sequences from Korean specimens and M. migitae from Japan (EU521643) were almost identical with 0.2% genetic distances. Lindstrom and Fredericq (2003) mentioned that the sequence divergences of rbcL within species of Porphyra from different populations differed by 0.1% in Porphyra fallax from British Columbia, Canada, to 0.9% in Porphyra pseudolanceolata and P. pseudolinearis from Alaska, USA. The sequence divergence between Korean and Japanese specimens fell into the intraspecific variation, showing the specimens to be conspecific. In the phylogenetic tree, Miuraea formed a clade with four genera, Pyropia, Wildemania, Boreophyllum, and Lysithea, without supporting value. These phylogenetic relationships were different from two previous studies: one is that the genus Miuraea formed a basal node of the order Bangiales except for two filamentous genera Dione and Minerva (Sutherland et al. 2011), in the other is that the genus Miuraea formed a sister clade composed of Boreophyllum, Fuscifolium, Pyropia, Wildemenia, and an undescribed filamentous genus (Sánchez et al. 2014). In addition, the taxonomic confusion still remained on the phylogenetic relationships of three genera, Porphyra, Bangia, and Clymene, even though they had moderate support value in the rbcL tree (Sutherland et al. 2011; Sánchez et al. 2014).
The new record of M. migitae from Korea extends the distribution range of this species previously restricted to only two localities, Osaka and Nagasaki in Japan (Guiry and Guiry 2016). Udo of Jeju Island is distant from the type locality of M. migitae, Osaka, Japan. Hwang (2008) mentioned that Udo had been connected with Osaka by shipping previously (before 1945). Although the origin of this species is unclear, this report indicated that this species might be occurred in Korea prior to that time. Lack of the records on the distribution of M. migitae from Udo before our finding might be due to the difficulty of collecting in its subtidal habitat where there are swift currents. Since the foliose thallus of M. migitae is growing up to 28 cm as the maximum length and producing asexual reproductive cells during summer season (Kikuchi et al. 2010), it can be a potential genetic resource for plant breeding to increase the production in seaweed aquaculture.
Conclusions
In conclusion, we collected the foliose Bangiales specimens from the subtidal zone of Udo, Jeju Island. Based on molecular and morphological analyses, these specimens were identified as M. migitae, previously considered endemic in Japan. This is a new record for Korea and indicates that collections from the subtidal zone are necessary to increase understanding of the biodiversity of seaweeds including potential genetic resources in Korea. In further studies, we are performing morphological and molecular investigations of the order Bangiales to understand the accurate phylogenetic relationships.