





Background
Insulin-like growth factor (IGF) plays important roles in regulating growth, metabolism, development, reproduction, osmoregulation, and the immune response (Li et al. 2012). IGF-I is a growth indicator in fish (Vera Cruz et al. 2006) and induces biological responses with specific receptors in target tissues. In addition, IGF-I and IGF-II play important roles in early embryonic development and growth in mammals and fish (Hardy and Spanos 2002; Chandra et al. 2011). Mammalian embryos require more time for differentiation and growth than do fish embryos, and they depend on various signal pathways generated by growth factors during pre-implantation (Riley et al. 2005). In fish, the expression of IGF-I and IGF-II mRNAs depended on the embryonic stage and was detected in different organs and at developmental stages (Xu et al. 2015; Yuan and Hong 2017).
The olive flounder Paralichthys olivaceus is an important economic marine aquaculture fish in Korea. In particular, early seed production, which is controlled by fertilization and hatching rate, is vital for aquaculture success. Therefore, to study the relationships between normal expression of growth factors, such as IGF-I and IGF-II, and normal development and embryo growth of the olive flounder, we investigated the involvement of IGFs and protein variation during embryonic development.
Methods
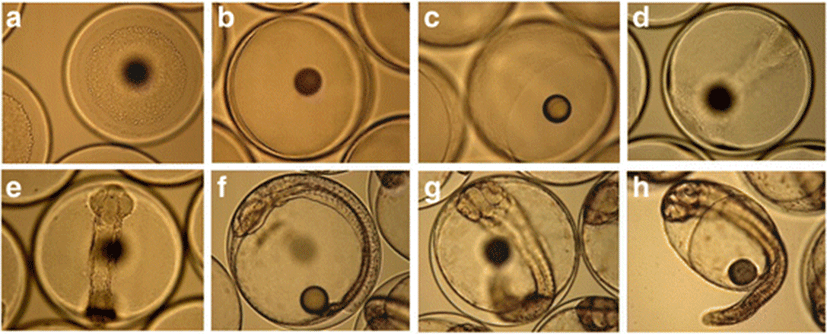
Embryos were obtained from the Korean Fish Breeding Institute, Jeju-Si, Jeju Special Self-Governing Province, Korea. They were maintained in 50-L square tanks (n = 3 pools) at 18 ± 0.5 °C. Samples were collected at the blastula (12 hours post-fertilization [hpf]), early gastrula (17 hpf), gastrula (21 hpf), cephalization (22 hpf), early cranial regionalization (27 hpf), late cranial regionalization (34 hpf), and tail lift (42 hpf) stages; stage classification followed Martinez and Bolker (2003) (Fig. 1). Samples (n = 50) were immediately frozen in liquid nitrogen and stored at − 75 °C until use.
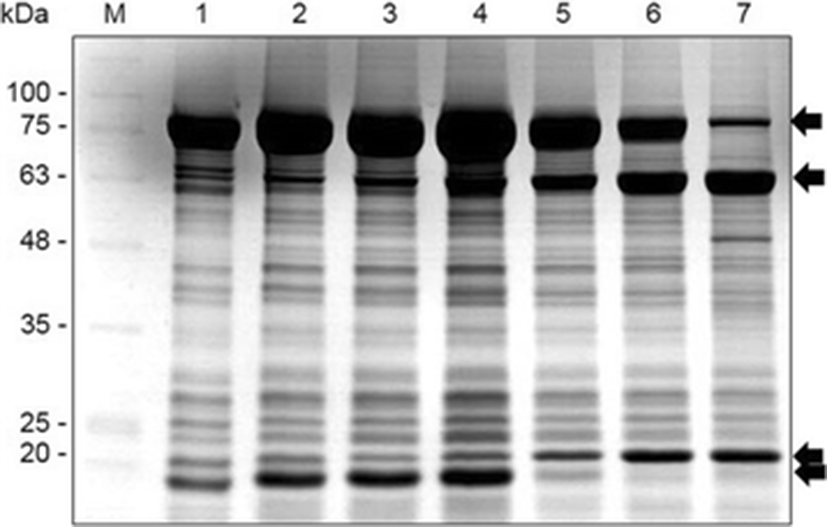
We investigated protein changes during embryonic development using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). Specific proteins were confirmed using electrospray ionization quadrupole time-of-flight mass spectrometry/mass spectrometry (ABI, USA), according to a previously described method (Choi et al. 2015). Proteins were identified via the UniProt Knowledgebase (http://www.uniprot.org/uniprot) using the MASCOT program (Matrixscience, London, UK).
To determine IGF involvement in embryo development, an immunoblot assay was performed. Embryos (n = 50) were homogenized in radio immunoprecipitation lysis buffer (50 mM Tris, 1 mM ethylene glycol-bis (2-aminoethylether)-N,N,N′,N′-tetraacetic acid, 150 mM NaCl, 1% NP-40, and 0.25% Na-deoxycholate) containing protease inhibitors (1 μg mL−1 aprotinin, 1 μg mL−1 leupeptin, 1 μg mL−1 pepstatin, 1 mM Na3VO4, 1 mM NaF, and 1 mM phenylmethanesulfonyl fluoride). Samples were centrifuged at 12,000 rpm for 10 min, and the supernatant was collected. Protein concentration was measured using the BCA Protein Assay Kit (Pierce Biotechnology, Rockford, IL, USA). Proteins (30–50 μg) were separated by 8–15% SDS-PAGE and transferred to polyvinyl difluoride membranes (Millipore, Billerica, MA, USA). The membranes were blocked with TBS-T (10 mM Tris–HCl, 150 mM NaCl [pH 7.5], and 0.1% Tween 20) containing 1% bovine serum albumin and incubated with primary antibody (1:1000 in TBS-T) on a rocker overnight at 4 °C. Primary antibodies used for immunoblotting included polyclonal rabbit anti-IGF-I, anti-IGF-II, anti-IGF-IR ß-subunit, anti-RAS, anti-GRB2, anti-p-AKT1/2/3, anti-mitogen-activated protein kinase kinase (MEK), monoclonal mouse p-Tyr (PY99), anti-extracellular signal-regulated kinase (ERK) 1/2, and anti-p-ERK (Santa Cruz Biotechnology, Santa Cruz, CA, USA). After washing in TBS-T, membranes were incubated for 1 h at room temperature with horseradish peroxidase (HRP)-conjugated goat anti-mouse immunoglobulin G (IgG, sc-2031; Santa Cruz Biotechnology) or HRP-conjugated goat anti-rabbit IgG (A-0545; Sigma-Aldrich, St. Louis, MO, USA) diluted 1:10,000 in TBS-T. Monoclonal mouse anti-ß-actin (C4) antibody (sc-47778; Santa Cruz Biotechnology) was used as a control. Protein was detected using enhanced chemiluminescence detection reagents (Advansta, Menlo Park, CA, USA) and visualized using the GeneSys imaging system (SynGene Synoptics, Ltd., London, UK).
Results
Protein expression depended on embryonic stage, and marked variations in protein expression were detected at four stages (Fig. 2, arrows). These proteins were identified as belonging to the vitellogenin 2 family (Accession no.: tr|Q5KQQ7|Q5KQQ7_PAROL, http://www.uniprot.org/uniprot/Q5KQQ7) by comparing its probable amino acid sequence with those in the NCBI and UniProt protein databases.
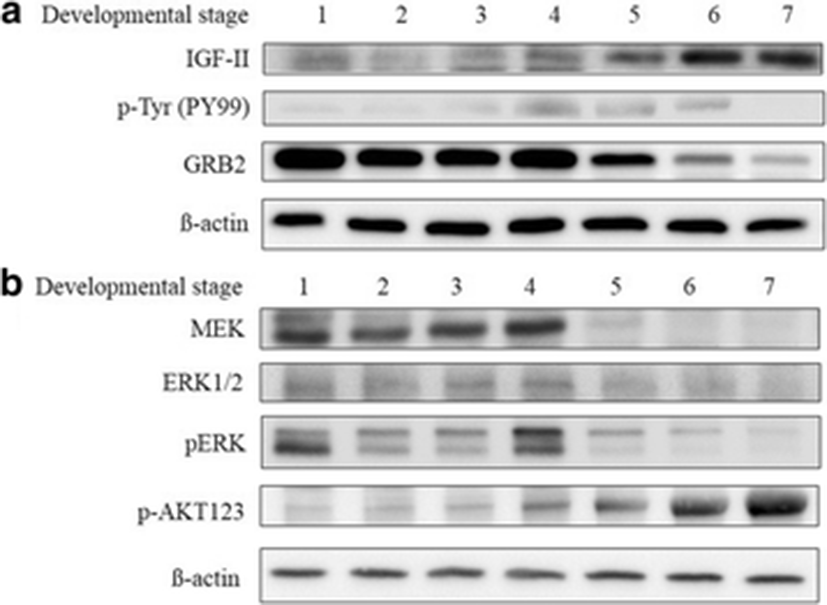
In this study, the IGF-IR β-subunit was not detected in all developmental stages. To determine whether the IGF-IR β-subunit was activated, tyrosine phosphorylation (PY99) was assessed using immunoblot analysis. Expression of PY99 throughout egg development was observed. IGF-II expression increased significantly as egg development progressed (Fig. 3a). These results suggest that binding of IGF-II to IGF-IR stimulates egg development. Docking of GRB2 to the phosphorylated IGF-IR β-subunit leads to ERK phosphorylation through the Ras/MEK axis (Fig. 3b). The expression of p-Akt increased during development (Fig. 3b).
Discussion
IGF signaling has been proposed to be a major factor in the growth and development of vertebrates (Duan and Xu 2005; Schlueter et al. 2007). The effects of IGFs on embryonic growth and development in fish have been examined in zebrafish (Schlueter et al. 2007; Zou et al. 2009) and rainbow trout (Li et al. 2010). Normal embryonic development and growth depend on a combination of internal and aqua-environmental factors (Choi and Chang 2003). The embryo is free floating, with no direct cell-to-cell contact, lacks a blood supply, and is exposed to a dynamic fluid environment (Hardy and Spanos, 2002; Riley et al. 2005). Therefore, the embryo depends on autocrine and paracrine growth factors to support its growth and development (Riley et al. 2005). The IGF-IGF receptor-IGF-binding protein system is a complex regulatory system that is essential for normal growth and development (Duan, 1997), and autocrine/paracrine IGF-I is the main determinant of postnatal body growth (Ohlsson et al., 2000). Also, the structure of the fish IGF-IR is very similar to that of mammals with an overall amino acid sequence identity of 72% (Duan 1997). These findings indicate that the IGF-IR is highly conserved both structurally and functionally in teleost fish (Choi et al. 2014). IGF-IR mediates IGF-I and IGF-II action in prenatal growth, and IGF-II action in postnatal growth, in mammals (Santos et al. 2008). Autocrine/paracrine roles of IGF-II in the differentiation, maintenance, and regeneration of central neurons and in the maturation of oocytes and follicle development in fish have been reported (Caelers et al. 2004). In this study, the expression of phosphotyrosine was examined, and IGF-II was found to be expressed after the cephalization stage, leading to the differentiation of such organs as the optic vesicles. IGF-IR activation leads to downstream activation of the RAS/MEK/ERK pathway and the Akt pathway. RAS/MEK/ERK is a key signaling pathway in cell proliferation and differentiation (Fuentes et al. 2011). Akt activation modulates the function of numerous downstream substrates involved in the regulation of cell survival and growth (Manning and Cantley 2007). In the present study, promotion of embryo growth occurred through the activation of ERK and Akt.
The expression of IGF-IR-related signaling is associated with changes in the expression of the vitellogenin 2 family. Oviparous teleost eggs accumulate a large amount of yolk protein as a nutrient store required by developing embryos (Matsubara et al. 1999). During vitellogenesis, yolk precursor proteins (vitellogenin [Vg]) are synthesized hepatically and regulated by estrogen (Matsubara et al. 1999). The presence of two types of Vg molecule (Vg A and Vg B) was reported in the barfin flounder, and these molecules function during oocyte maturation and embryonic development (Matsubara et al. 1999). In this study, changes in vitellogenin 2 family expression were observed during embryonic development. The molecular weights of native Vg were estimated to be 550,000 in Platichthys flesus (Emmersen and Petersen 1976) and 530,000 in Scophthalmus maximus (Silversand and Haux 1989). In this study, higher-molecular-weight Vgs may have degraded over time and become involved in embryonic development. Further studies regarding the association between the function of cleaved Vgs and embryonic developments are necessary.
Conclusions
IGF-II and IGF-IR are expressed during embryonic development and are associated with the IGF-IR signaling pathway. The IGF signaling cascade may be involved in developmental regulation. Further study is needed to determine the relationship between Vg and the IGF system during embryonic development of the olive flounder.