






Introduction
Starfish is an abundant group of marine invertebrates that compromises approximately 1900 species. It belongs to the phylum Echinodermata, which includes five classes: Crinoidea, Ophiuroidea, Asteroidea, Echinoidea, and Holothuroidea (Mah et al., 2012; Martindale et al., 1998; Pawson, 2007). These echinoderms share a significant feature in common: a unique type of symmetry called radial symmetry. Radial symmetry is complexly superimposed on the remains of the bilateral symmetry from the larval stages (Davydov et al., 1990; Ji et al., 2012; Martindale et al., 1998). Starfish also has a unique digestive system enabling them to efficiently digest organism that are several times larger than its own size. Starfish’s cardiac stomach extends through its mouth to digest food outside the body. Then, partially digested food is passed to the pyloric stomach located in the interior of starfish’s body, and digestion is continued in the organ. The digested nutrient is then absorbed in digestive glands called pyloric caeca that are located in starfish’s arms. These pyloric caeca are lined with zymogen cells that secrete enzymes and absorb essential vitamins from food (Anderson, 1953, 1959, 1979).
Neuropeptides are biologically active compounds that participate in various mechanisms such as the regulation of metabolism, memory, pain perception, maintenance of homeostasis, and immune processes. Extracellular modification is a unique feature of neuropeptides by which peptides are cleaved through the peptidases (Russo, 2017). Neuropeptides are cleaved from precursor proteins by sequential hydrolytic degradation. Thus, it is possible to have numerous neuropeptides formed from one precursor protein. These extracellular cleavages can either inactivate biological activity or increase the peptide’s affinity for a particular receptor decreasing its affinity for another receptor (Hook et al., 2008). Some neuropeptides are modified by C-terminal amidation, which is required for biological activity (Merkler, 1994). Neuropeptides have been known as chemical signals for over 40 years. Some neuropeptides function as mediators carrying out the direct transmission of a nerve impulse, and other neuropeptides function as neuromodulators affecting the metabolism of the cell (Russo, 2017).
To date, 40 neuropeptide precursors have been discovered in starfish Asterias rubens using the transcriptome data. However, the functional activity of the majority is unknown (Semmens et al., 2016). The first isolated neuropeptides with a wide range of bioactivity within phylum Echinodermata were SALMFamide-1 (S1) and SALMFamide-2 (S2). Both neuropeptides were purified through a radioimmunoassay from two starfish species A. rubens and Asterias forbesi. These peptides induce relaxation of cardiac stomach and tube foot preparations in vitro and inversion of cardiac stomach in vivo (Elphick et al., 1991, 1995, 2013; Melarange et al., 1999). Other well-known neuropeptides are NG family peptides. These are widely distributed neurophysin-associated class of peptides within phylum Echinodermata. Whereas SALMFamide induces cardiac stomach eversion and relaxation, the NG neuropeptides induce cardiac stomach retraction in A. rubens (Elphick, 2010). Moreover, NGFFYamide causes the relaxation of the apical muscle and the contraction of the tube feet and significantly decreases the velocity of locomotor activity (Semmens et al., 2013; Tinoco et al., 2018). Asterotocin is a VP/OT-type neuropeptide that causes, like SALMFamide, the relaxation of cardiac stomach, but with more potent activity than SALMFamide-2; asterotocin also causes cardiac stomach inversion in A. rubens (Gruber, 2014; Odekunle et al., 2019).
Another muscle relaxant neuropeptide designated as starfish myorelaxant peptides (SMP) was purified from the whole body of the P. pectinifera and was identified as a member of the PP/OK- type neuropeptide family (Kim et al., 2016). Pedal peptide (PP) and orcokinin (OK) are two structurally related neuropeptides discovered from two protostomean phyla (phylum Mollusca and phylum Arthropoda). PP was identified from pedal ganglia of mollusk Aplysia californica and OK was identified from the crayfish Orconectes limosus with a myotropic effect (Stangier et al., 1992; Wang et al., 2021). In Echinodermata (deuterostome), PP/OK neuropeptides were discovered using the transcriptome data of sea urchin Strongylocentrotus purpuratus and sea cucumber Apostichopus japonicas (Rowe & Elphick, 2012). The discovery of PP/OK-type peptides including SMP in the deuterostomes suggests that PP/OK-type neuropeptides are a bilaterian neuropeptide family (Kim et al., 2016). PP/OK-type neuropeptides have a broad spectrum of relaxation/contraction activity in mollusks and arthropods. Meanwhile, the stimulatory effect was the main physiological function of these peptides (Lin, 2017). The investigation on the physiological role of SMP revealed that SMP had muscle-relaxation effect, which was contrary to contractile effect of PP/OK neuropeptides (Kim et al., 2018). The cDNA cloning of SMP revealed that the precursor protein contains nineteen peptides that could be grouped into 4 isotypes: SMPa, SMPb, SMPc, SMPd. The main isotype, SMPa, had 12 copies while SMPb had 5 copies, and SMPc and SMPd each had one copy (Kim et al., 2016).
The investigation on the physiological role of SMP using three muscle preparations (cardiac stomach, tube feet, and apical muscle) revealed relaxation activity in P. pectinifera. SMP affects feeding behavior and the locomotion activity of starfish. SMP has a more potent relaxation activity than SALMFamide-1 and SALMFamide-2, had more potent relaxation activity (Kim et al., 2016). The primary structures of SMPs are as follows: SMPa (FGKGGAYDPLSAGFTD), SMPb (FGMGGAYDPLSAGFTD), SMPc (FGMGGAYDPLSAGFTE), and SMPd (GFLHGPDDPLSTSFVDGD). The comparison of SMP isotypes’ bioactivity revealed that SMPa was the most potent relaxant on apical muscle, cardiac stomach, and tube feet preparations (Kim et al., 2018). Here we report the isolation of a SMP isotype (SMPb) from the starfish P. pectinifera.
Materials and Methods
Live specimens of the starfish, Patiria pectinifera, were collected in Busan, Korea. The specimens were maintained in a recirculating seawater system at 13°C until use. Approval by the local institution/ethics committee was not required for this work because experimental work on starfish is not subject to regulation.
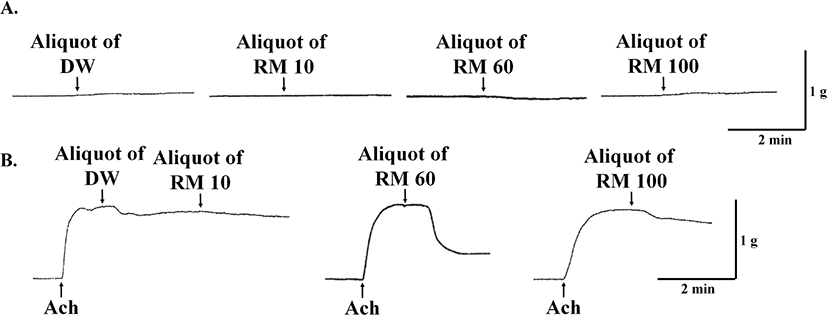
Starfish (P. pectinifera) were cut open using scissors, and the pyloric caeca was collected using forceps. The collected tissues were then boiled in 4 volume of water for 5 min and cooled on ice. Then, the boiled tissues were homogenized and glacial acetic acid was added to yield a final concentration of 5% acetic acid. The homogenate was then centrifuged (7,000×g, 20 min, 4°C). The pellet was re-extracted twice in 5% glacial acetic acid with the same extraction method. The supernatant was pooled and concentrated using a rotary evaporator. The concentrated extract was diluted with 4 volumes of methanol and then was centrifuged (7,000×g, 20 min, 4°C) to remove the precipitates. The supernatant was evaporated and then 6 volumes of ethanol was added. The precipitates were removed through centrifugation (7,000×g, 20 min, 4°C), and the supernatant was concentrated using a rotary evaporator. After concentration, the 0.1 volume of 0.1 M hydrochloric acid was added. The precipitates were removed again by centrifugation (7,000×g, 20 min, 4°C), and the supernatant was applied to a C18 cartridge (Sep-Pak C18 20 cc Vac Cartridge, Waters, Milford, MA, USA). The cartridge was washed with water/0.1% trifluoroacetic acid (TFA) and retained materials (RM) were then eluted with 10% methanol/0.1% TFA, 60% methanol/0.1% TFA, and 100%methanol/0.1% TFA. The obtained materials were named as RM10, RM60, and RM100, respectively. The eluates were evaporated and its biological activity on the apical muscle of P. pectinifera was investigated, as described below in the methods section for in vitro bioassay.
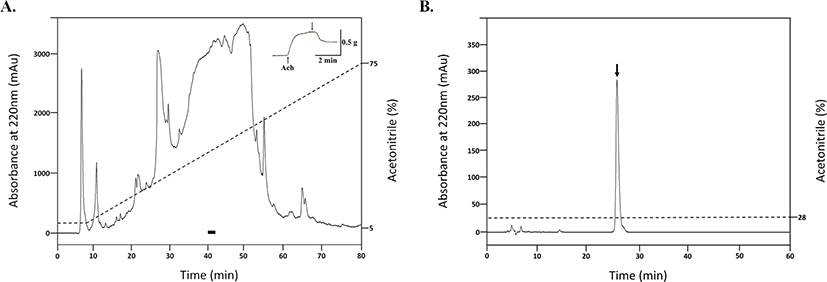
The obtained eluents were screened through the bioassay system described in the ‘in vitro bioassay’ section using 1/2,000 of the total eluents. The fraction (RM60) with the most potent relaxation activity was used for the purification. The RM60 was lyophilized, dissolved in water, and then applied to a reversed phase column (Capcellpak C18, 10 × 250 mm; Shisheido, Tokyo, Japan), which was eluted with a linear gradient of 5%–85% acetonitrile/0.1% TFA at flow rate of 2 mL/min for 80 min. Absorbance peaks were monitored at 220 nm and fractions were collected every 2 min by a fraction collector. The bioactive fractions, which were eluted between 40 to 42 min, were pooled, lyophilized, and then subjected to a cation exchange column (TSKgel SP-5PW, 7.5 × 7.5 mm; Tosho, Tokyo, Japan) with a linear gradient of 0–1 M sodium chloride in 20 mM phosphate buffer (pH 6.0) for 90 min at flow rate of 1 mL/min. A fraction eluted in 0 M sodium chloride caused relaxation of the apical muscle from P. pectinifera, which was then used for further purification steps.
An active fraction, which was eluted between 3 to 4 min, was pooled, lyophilized, and then applied to a second reverse phase column (Capcellpak C18, 4.6 × 250 mm; Shisheido). Elution was performed with a linear gradient of 5%–65% acetonitrile/0.1% TFA at a flow rate of 1 mL/min for 60 min, and each peak was collected manually. The peaks with relaxation activity were pooled, lyophilized, and was subjected to an anion exchange column (Mono Q HR 5/5, 50 × 5 mm; Pharmacia Biotech, Uppsala, Sweden) with a linear gradient of 0–1 M sodium chloride in 20 mM Tris-HCl (pH 8.0) at a flow rate of 1 mL/min for 50 min. The active peak eluted approximately 0.6 M sodium chloride was applied to further reverse phase column (Capcellpak C18, 4.6 × 250 mm) with a linear gradient of 25%–45% acetonitrile/0.1% TFA at flow rate of 1 mL/min for 40 min. To perform the final purification, the obtained peak was applied to the same column used in the previous step. Elution was performed with an isocratic elution with 28% acetonitrile/0.1% TFA at a flow rate of 0.5 mL/min.
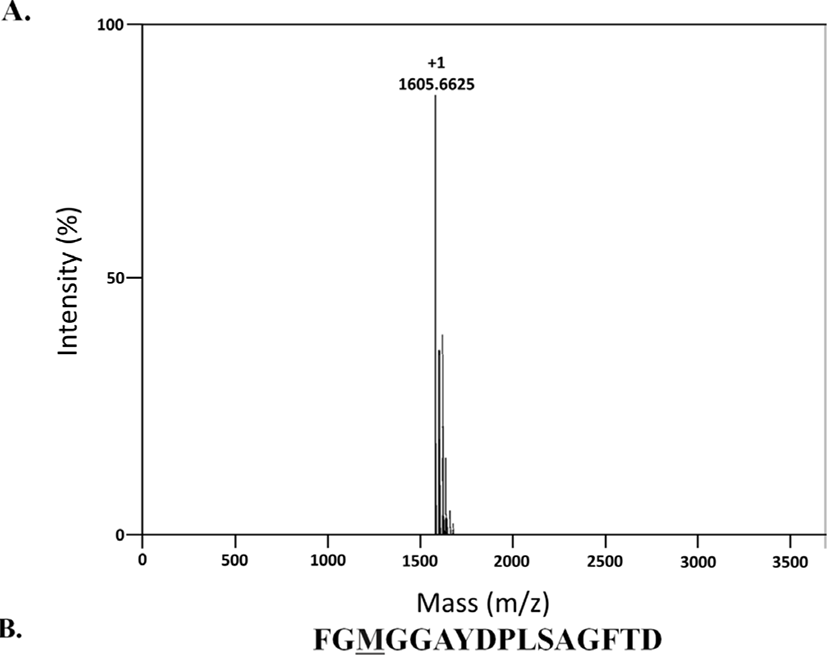
To determine the molecular mass of the purified peptide, liquid chromatography coupled with mass spectrometry (maXisT SHD Ultra-High Resolution Q-TOF, Burker, Germany) was used. To analyze the amino acid sequence of the purified peptide, an automated N-terminal amino acid gas-phase sequencer (PPSQ-1; Shimadzu, Kyoto, Japan) was used.
To determine the obtained fractions’ activities throughout the purification steps, a bioassay system measuring relaxation effect on the pre-contracted apical muscle preparation of P. pectinifera was used. Apical muscle with approximately 10.0 mm in length was carefully cut from the center of the starfish’s five arms. The muscle preparation was placed vertically in a polypropylene organ bath with aeration placed on the bottom side. The both ends of the cut muscle were tied by cotton threads; a loop was made with the cotton thread on the bottom end of the muscle, and the loop was connected to the bottom end of the bath through a silver wire. The top end of the muscle was attached to a force displacement transducer (Type 45196A; NEC-Sanei Instrument, Tokyo, Japan) with the cotton thread. The contraction or relaxation response of the muscle preparation was monitored and recorded by a recorder (WR7300; Graphtec, Yokohama, Japan) through an amplifier (AS1302; NEC-Sanei Instrument). The resting tension was adjusted to 1.5 g, and the muscle preparations were stabilized and equilibrated for 1 hour in advance of bioassay; during this time, a fresh replacement of artificial sea water (ASW) in the organ bath was carried out every 15 minutes. Pre-contraction of muscle preparations was performed by applying acetylcholine (Ach; final concentration of 5 × 10−6 M), and after equilibration, the test samples were applied to measure the muscle responses. The each obtained fraction aliquot was lyophilized, dissolved with 50 μL of ASW, and added into the organ bath. The bioassay was performed with 55 mM Mg2+ ASW with following composition: (mM) NaCl 445, KCl 10, CaCl2 · 2H2O 10, MgCl2 · 6H2O 55, Glucose 10, Tris-HCl 10 (pH 7.8).
Results
To purify neuropeptides from pyloric caeca of P. pectinifera, stepwise eluents (D.W., RM10, RM60, RM100) from the Sep-Pak C18 solid-phase extraction cartridge were screened for contractile and relaxation responses on the apical muscle of P. pectinifera. The experiment revealed that the most potent relaxation activity was observed from RM60, which was utilized for the further purification steps (Fig. 1). RM60 was first applied to a reverse phase column (Fig. 2A). The active fractions between 40 to 42 min were pooled and subjected to a cation exchange column. Unbound materials eluted between 3 to 4 min showed relaxation activity; therefore, used for further purification. The fraction was subjected to a second reverse phase column, where fractions between 38 to 39 min exhibited relaxation activity, and this fraction was further purified using an anion exchange column. An active fraction with a potent relaxation activity was eluted at 0.6 M of sodium chloride, which was applied to the third reverse-phase column. In the final purification step, the obtained fraction was separated through the reverse-phase column. A single absorbance peak was isocratically eluted with 28% acetonitrile/0.1% TFA at 25.8 min (Fig. 2B). The molecular weight of the isolated compound determined with LC-MS was 1,605.6625 Da (Fig. 3A). Using the Edman degradation method, the purified peptide was identified as an isotype of SMP with the sequence Phe-Gly-Met-Gly-Gly-Ala-Tyr-Asp-Pro-Leu-Ser-Ala-Gly-Phe-Thr-Asp which was previously discovered through the cDNA cloning of a SMP precursor protein (Fig. 3B).
Discussion
In this study, we have confirmed the existence of SMPb, an isotype of SMPa, that have been previously discovered through the cDNA cloning of SMPa’s precursor protein (Kim et al., 2016). SMPb was isolated for the first time from the pyloric caeca of starfish P. pectinifera. The isolation of SMPb confirms that the cleavage of SMP isotypes from the precursor protein takes place and that the physiological activity as muscle relaxant of P. pectinifera muscle preparations was present. SMPb possesses relaxation activity on three muscle preparations (apical muscle, tube feet and cardiac stomach) from P. pectinifera. Comparison of SMPb’s activity with that of SMPa revealed that there is no significant difference in the potency of the neuropeptides on all three muscle preparations (Kim et al., 2018). This could be attributable to the little change in the amino acid sequences of SMPb and SMPa; these two peptides are different merely at one position. The third position in SMPb’s sequence is a sulfur-carrying methionine residue whereas SMPa’s sequence has a basic residue, lysine, at the same position.
The alignment of SMPa, SMPb, SMPc from various starfish species revealed the existence of two conserved regions (Fig. 4). The first conserved region is composed of Phe-Gly at position 1 and 2 and is shared among all SMP peptides except for ArPPLN1a from A. rubens. The second region is composed of GAYDPLSAGF, which is known as the core bioactive region (Kim et al., 2018). The alignment of SMPs revealed the substitution of an amino acid at position 7 (from Tyr to Phe) in both A. rubens and Asterias amurensis. The highest homology of SMPs from P. pectinifera was observed with the SMPs from Acanthaster planci, which were identified through in silico analysis (Smith et al., 2017). Moreover, both SMPa and SMPb from P. pectinifera are 100% identical to SMPa and [Met3]-SMPa from A. planci, respectively. This could be attributable to the evolutionary divergence of the two orders: Valvitida and Forcipulatida. P. pectinifera and A. planci belong to Valvitida whereas A. rubens and A. amurensis belong to Forcipulatida.
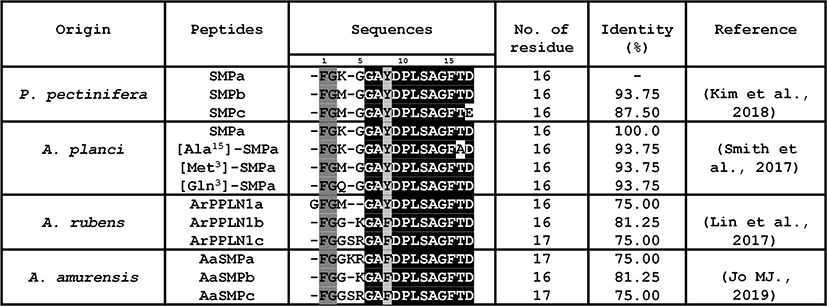
A previous study on the tissue distribution of SMP transcript has revealed that the highest expression was observed in radial nerve cord. The expression levels in digestive organs such as pyloric caeca, pyloric stomach, and cardiac stomach was the lowest (Kim et al., 2018). The presence of myorelaxant peptides in pyloric caeca might be attributable to the inclusion of the thin and thick myofilament bundles in the current-producing cells (Martinez, 1989). The wall of pyloric caeca consists of three layers: an outer mesothelial layer, a middle layer containing connective, muscular, and nervous tissues, and an inner digestive epithelium layer (Chia, 1969). The digestive epithelium contains cells such as, secretory and endocrine cells, and the special current-producing cells that possess bundles of myofilaments (Martinez, 1989). Interestingly, SMPb analog merely comprising fifteen residues, without the sixteenth aspartic acid residue, was also isolated from the pyloric caeca of P. pectinifera. The molecular weight of the isolated compound determined with LC-MS was 1,490.6630 Da (data not shown). The investigation of the synthetic SMPb’s bioactivity revealed that SMPb exhibited relaxation effect on cardiac stomach, tube feet, and apical muscle (Kim et al., 2018). Although there had not been reports of SMPs’ other activities, it is possible that SMPs could induce relaxation of smooth muscles from other animals due to the multifunctional nature of neuropeptides. Thus, further investigation on the bioactivities of SMP-related peptides may lead to the discovery of muscle relaxants from other Echinoderms.