






Introduction
Spiny lobsters of the genus Panulirus are globally renowned for their exquisite flavor and economic significance in the seafood market. Of the 19 recognized species worldwide, six are found in South Korean waters: Panulirus japonicus, Panulirus penicillatus, Panulirus stimpsoni, Panulirus versicolor, Panulirus homarushomarus, and Panulirus longipes (Holthuis, 1991). Among these, the present study focused on P. stimpsoni and P. japonicus, as these species inhabit Jeju Island, South Korea (Hettiarachchi et al., 2022). Research on the biology, ecology, and distribution of these species is limited in South Korea, largely because they are not considered major fishery commodities on the peninsula. This lack of information has hindered efforts to explore the potential of spiny lobsters as a viable South Korean fishery stock. Moreover, the management and conservation of these species are further complicated by the limited ecological and biological data available.
The coastal geomorphology of Jeju Island, characterized by rocky substrates, provides an optimal habitat for spiny lobsters (Holthuis, 1991). This unique environment supports the life cycle of Panulirus lobsters, which includes two distinct phases: a planktonic larval stage and a benthic post-larval stage (Caputi et al., 1995). Notably, P. japonicus undergoes an ontogenetic niche shift, where post-larvae settle in habitats distinct from those occupied by adults (Briones-Fourzán & Lozano-Álvarez, 2013). In terms of feeding behavior, Panulirus lobsters, including those from Jeju, are opportunistic omnivores, primarily foraging during nocturnal hours. As they grow, their foraging range expands, reflecting an adaptable feeding strategy. Despite the high demand for lobsters in countries like the United States, Japan, and China, the potential of P. japonicus and P. stimpsoni as fishery stocks in South Korea remains largely unexamined. The limited understanding of their reproductive biology, population dynamics, and ecology presents substantial challenges to effective resource management.
In this study, we aimed to address these gaps by providing essential information on the population structure of P. japonicus and P. stimpsoni. The primary objectives were to investigate the relationship between carapace length and body weight, assess the condition factors of wild populations, and clarify the stages of egg development in palinurid lobsters. Ultimately, this study seeks to offer valuable insights into the sustainable utilization and management of these marine resources, unlocking their untapped potential for the fishery sector in South Korea.
Materials and Methods
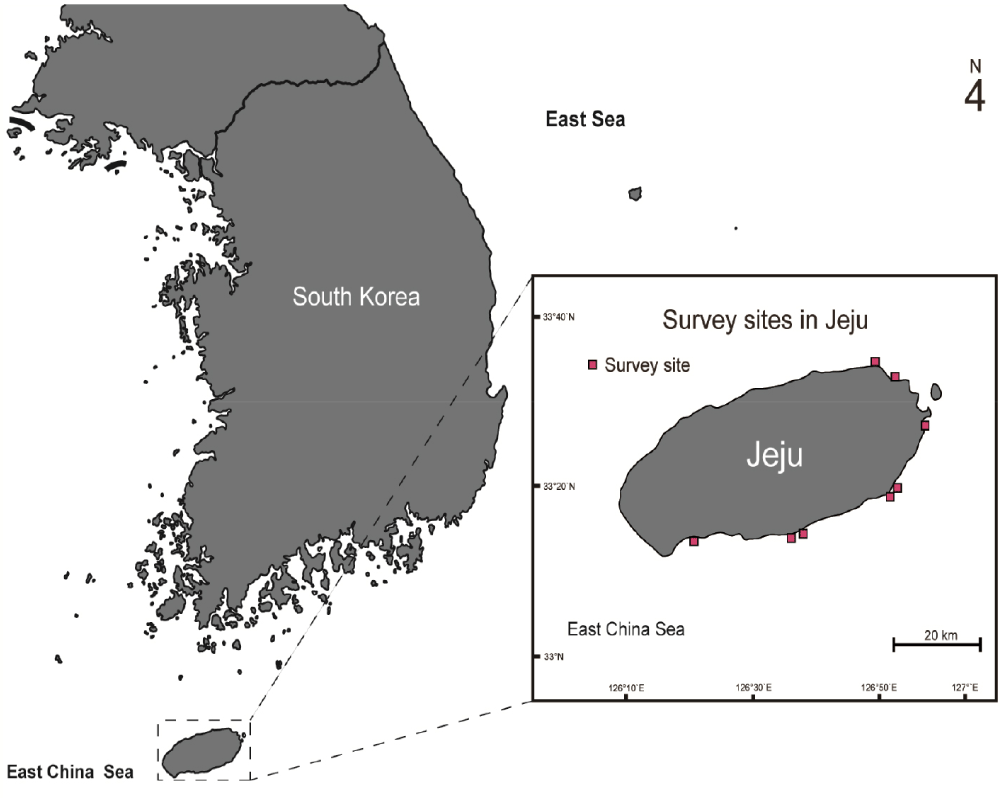
Adult live spiny lobsters (P. stimpsoni female n = 16, male n = 9; P. japonicus female n = 13, male n = 14) were collected from eight sampling sites along the southern and southeastern coasts of Jeju Island, South Korea, by scuba diving (Fig. 1) between January and December 2020. The captured lobsters were transported to circular indoor tanks.
The carapace length (mm) and body weight (g) were recorded separately. The carapace length (mm) was measured using a ruler to the nearest millimeter, from the orbital notch to the hind edge of the carapace along the mid-dorsal line (Farmer, 1975, cited by Senevirathna et al., 2014). All lobsters were transferred to acrylonitrile butadiene styrene tanks (20 tons) with a constant circulation of seawater and maintained at a temperature of 23 ± 1°C under natural light. The lobsters were fed a commercially formulated powdered feed daily at 16:00 (Heukja, Kopec, Gimcheon, Korea). The tanks were provided with polyvinyl chloride shelters. Berried females were reared in separate tanks, and molting events were recorded by collecting exoskeletons. The developmental stages of the eggs of berried individuals were observed under a photomicroscope (Nikon ECLIPSE Ni, Tokyo, Japan) to identify the developmental stage and measure the mean egg diameter at each stage (n = 30/developmental stage). The embryonic development stages of P. polyphagus described by Ikhwanuddin et al. (2014) were used as a reference for identifying egg stages. The criteria for distinguishing egg development included egg color, eye spot, appendages, chromatophores, and yolk.
The relationship between carapace length and body weight of the two Panulirus lobsters was analyzed using the following formula (Ricker, 1975):
Where:
W, body weight (g),
L, carapace length (mm),
a, constant (intercept), and
b, slope or the allometric growth coefficient.
The condition factor was calculated based on species and sex using the subsequent formula (King, 2007). Pearson’s correlation coefficient was used to estimate the strength and pattern of relationships between the two variables.
where:
CF, condition factor,
W, actual body weight,
Wp, estimated body weight (derived from Wp = aLb).
Differences in the mean values of the condition factors for male and female individuals of the two species were determined using the non-parametric Mann-Whitney U test. The difference in mean egg diameter across the identified stages was determined using a one-way analysis of variance (ANOVA), followed by Tukey’s HSD test. All statistical analyses were conducted with a 95% confidence level using GraphPad Prism ver. 8.4.3 (https://www.graphpad.com/features).
Results
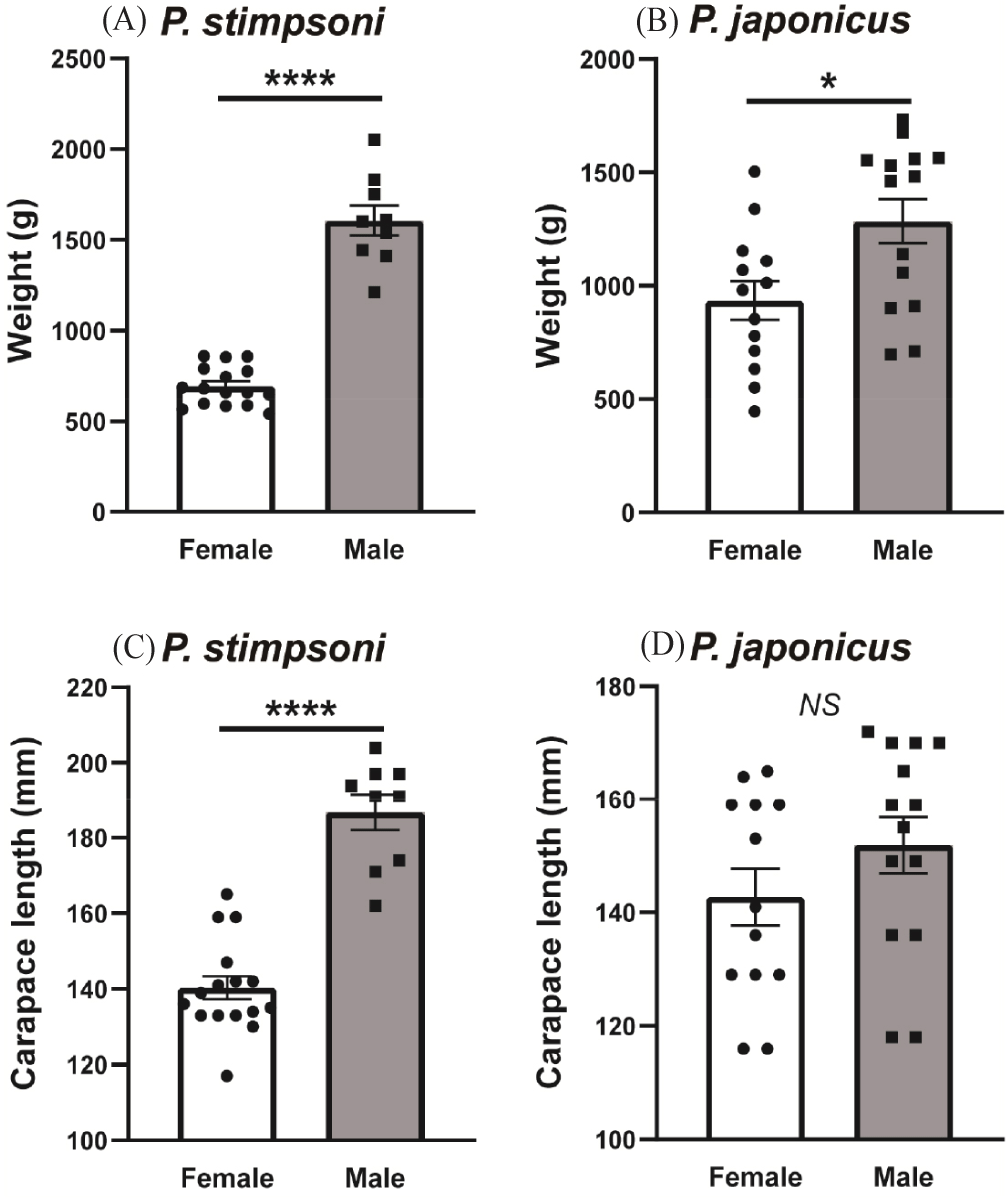
A total of 52 lobster specimens were captured during the study, comprising two species: P. japonicus and P. stimpsoni (Fig. 2). Among the sampled individuals, the male-to-female sex ratio was approximately 1:2 for P. stimpsoni and 1:1 for P. japonicus (Fig. 2A and 2B). The mean weight of male P. stimpsoni was 1,605.22 ± 82.89 g, whereas females weighed 692.00 ± 27.02 g on average. The observed mean weight difference between male and female P. stimpsoni was statistically significant (p < 0.05; Fig. 2A). For P. japonicus, the mean weight of the males and females was 1,284.07 ± 97.89 g and 934.23 ± 85.86 g, respectively (Fig. 2B). Regarding carapace length, for both P. stimpsoni and P. japonicus, males tended to have a longer carapace than that of females. For instance, male P. stimpsoni exhibited a mean carapace length of 186.78 ± 4.74 mm, whereas females showed a mean length of 140.31 ± 3.07 mm (Fig. 2C). Similarly, the mean carapace length for male P. japonicus was 151.86 ± 4.97 mm, whereas it was 142.69 ± 4.99 mm for females (Fig. 2D). Despite these differences, the variation in mean carapace length between males and females of both species was not statistically significant (p >0.05).
Both female and male individuals of P. japonicus and P. stimpsoni displayed a strong, significant correlation between carapace length and body weight (p < 0.05, R2 ≈ 1.0; Fig. 3).
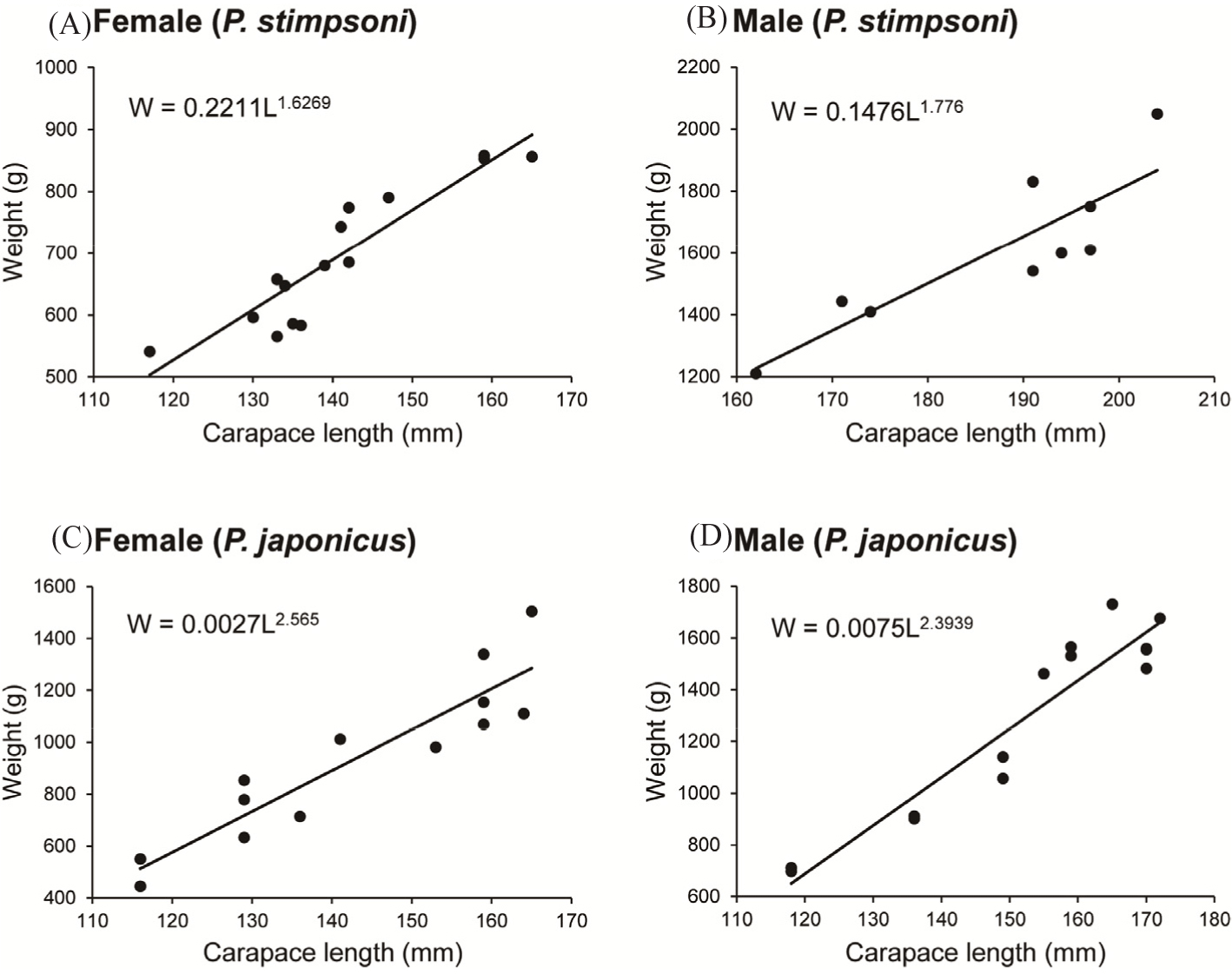
The allometric growth coefficients of both male and female individuals in the two species did not exceed 3.0, indicating that both female and male P. stimpsoni (Fig. 3A and 3B) and P. japonicus (Fig. 3C and 3D) exhibited negative allometric growth. The allometric growth coefficient was visibly higher in both male and female P. japonicus than in P. stimpsoni (Table 1).
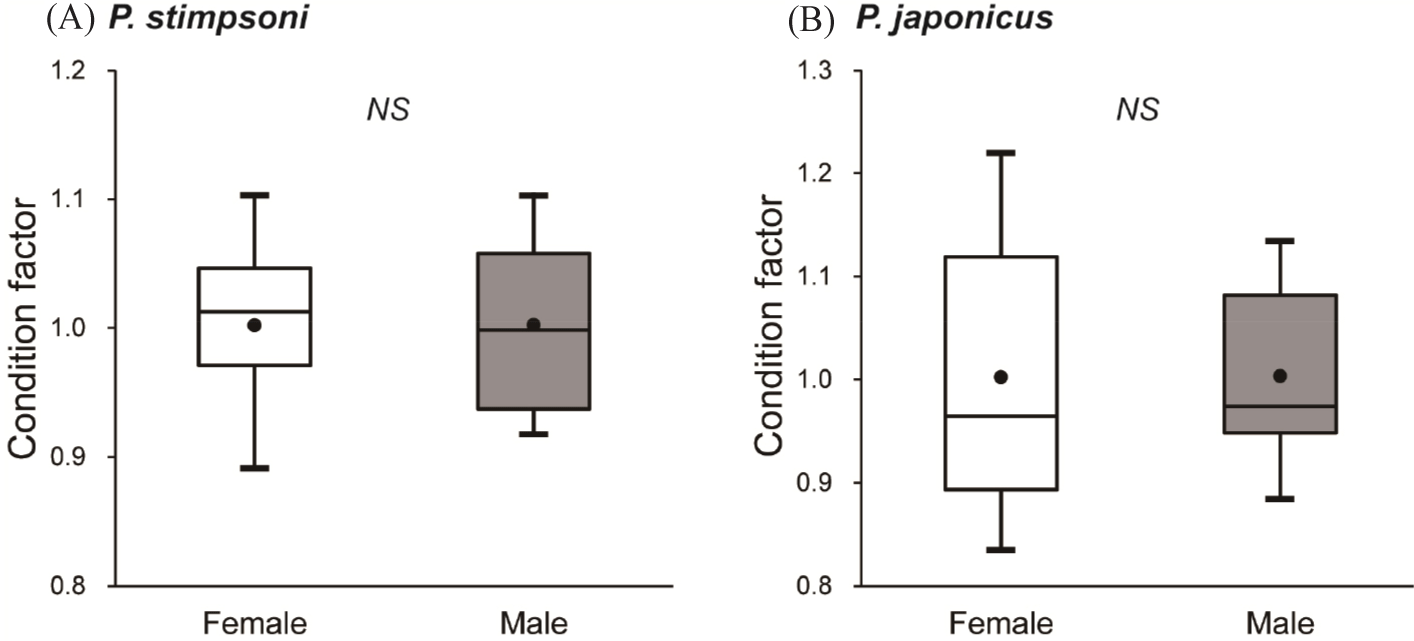
The average condition factor of male and female individuals from both lobster species was ≥ 1.00 (Fig. 4). The mean condition factor for both male and female P. stimpsoni was 1.00 ± 0.02, with no significant differences observed (Fig. 4A). In contrast, male P. japonicus exhibited a mean condition factor of 1.01 ± 0.02, and females displayed a similar mean condition factor of 1.01 ± 0.04. However, the condition factor of female P. japonicus was significantly higher than that of its male counterpart (Fig. 4B).
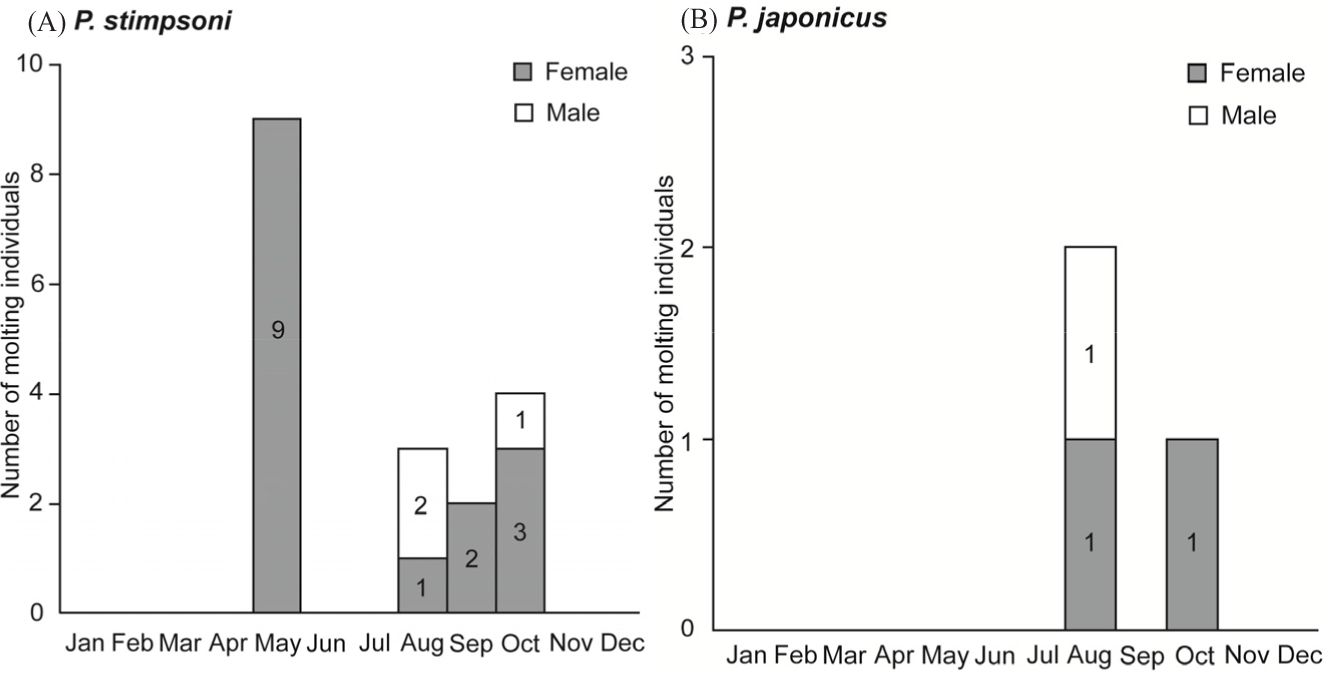
Molting was presumed to occur at night during the experiment. Two molting regimes were observed for females and one for males. Molting was predominantly observed in P. stimpsoni, whereas a few individuals of P. japonicus molted during the experimental period. Molting of P. stimpsoni occurred at two distinct time points: one in May and the other from August to October (Fig. 5A). Molting occurred less frequently in P. japonicus, and only three individuals molted during the experimental period (Fig. 5B).
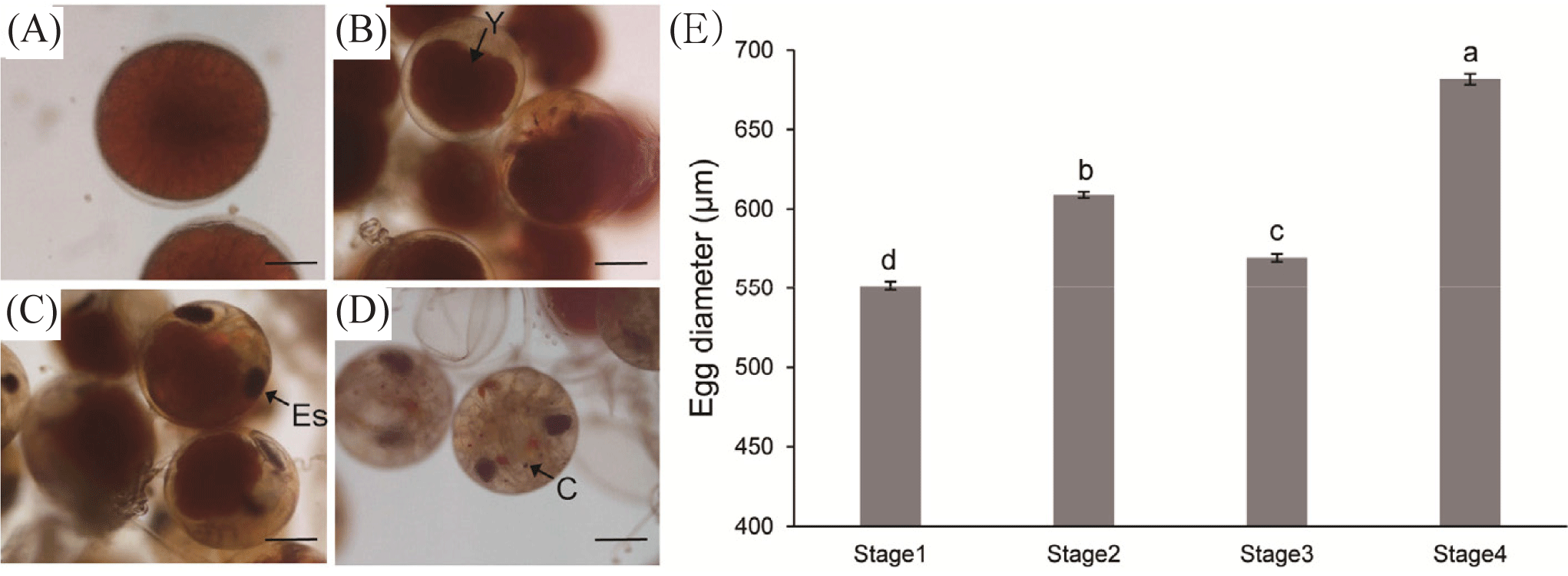
Four distinguishable egg stages were identified, with Stage 4 reached after approximately one month (Fig. 6). At Stage 1, eggs were dominated by a brick-red yolk (Fig. 6A). At Stage 2, approximately two weeks later, the yolk shrank, and appendages became visible (Fig. 6B). At Stage 3, approximately three weeks later, the yolk further shrunk, and eye spots were visible (Fig. 6C). At Stage 4, approximately one month later, the yolk was no longer visible, chromatophores emerged throughout the appendages, and the eye spot was larger and more complex (Fig. 6D). The mean egg diameter at each successive stage showed an increasing trend, except for Stage 3, where the mean egg diameter significantly reduced compared to Stage 2 (p < 0.05; Fig. 6E).
Discussion
In both P. japonicus and P. stimpsoni, the male lobsters were slightly larger than the female lobsters in terms of weight and length. Larger male lobsters may reflect the influence of mating success on their size, which is consistent with the findings of Butler IV et al. (2015). The sex ratio for captured P. japonicus was 1:1, while that for P. stimpsoni was 1:2. Lobster population size, structure, and sex ratios tend to vary with depth, season, and site geomorphology (MacDiarmid, 1991). However, the sex ratio observed in the present study may represent the population structure of the area or merely a sampling bias caused by the limited number of samples, as the number of specimens was restricted due to the absence of commercial spiny lobster fisheries on Jeju Island and the excessive effort and cost involved in sampling. Three major reasons for the skewed sex ratio in spiny lobster samples are: 1) sex-specific catchability, 2) sex-specific natural and/or fishing mortality, and 3) sex-specific movement patterns (Jury et al., 2019). Furthermore, Koepper et al. (2021) identified bottom temperature and water depth, which impact their growth, reproduction, and survival, as determinants of sex ratios in lobsters. Although a 1:1 sex ratio is not necessarily required for a healthy population, skewed sex ratios may eventually influence mating dynamics, potentially affecting the sustainability of the population in the long run (Jury et al., 2019). Therefore, the sex ratio of broodstock must be considered for the sustainable management of these living resources.
According to Pearson correlation coefficients, both female and male individuals of P. japonicus and P. stimpsoni exhibited strong and statistically significant relationships between carapace length and body weight (p < 0.05; Fig. 3). Isometric growth occurs when the allometric growth coefficient (b) equals 3 (Senevirathna et al., 2014). When the allometric growth coefficient is greater than 3 (b > 3), this indicates positive allometric (hyperallometric) growth, meaning weight increases faster than length. Conversely, an allometric growth coefficient smaller than 3 (b < 3) reflects negative allometric (hypoallometric) growth, where length increases faster than weight (Bensahla-Talet et al., 2016). In this study, both male and female P. japonicus had higher allometric growth coefficients than P. stimpsoni. However, the allometric growth coefficient for female P. japonicus was slightly higher than that for males, while the opposite pattern was observed for P. stimpsoni. Allometry indicates how an organism changes its shape and proportions as it grows (MacCormack & Demont, 2003). Typically, this leads to differences in growth rates among body parts (Vogel, 1988), a phenomenon known as allometric growth. Previous studies suggest that the allometric growth coefficient of spiny lobsters varies due to factors such as age, sex, maturity stage, season, temperature, and food availability (Senevirathna et al., 2014). Regional differences in growth rates for the same species have also been noted (MacCormack & Demont, 2003). In artificially reared spiny lobsters, carapace length was found to increase with age (Hutchinson et al., 2024). However, in the case of common lobsters, studies have reported a weaker relationship between size and age. For lobsters reared in controlled environments, the growth rates among individuals exhibit less variability, allowing size to serve as a reliable indicator of age. In contrast, under natural conditions, growth rates are influenced by various environmental factors, making size an unsuitable proxy for age (Maxwell et al., 2007). As growth parameters vary between stocks, areas, and specimen numbers examined (Dulčić & Kraljević, 1996), further studies are necessary to better understand these variations and support future population management.
The condition factor is a metric used to assess individual health in a population or to compare the well-being of different populations (Situmorang et al., 2021). It indicates that individuals of a certain length but greater weight are in better condition than others. The relative condition factor employed in this study is appropriate for comparing conditions between different populations and species (Froese, 2006).
In this study, female P. japonicus exhibited a higher mean condition factor than male individuals. Further sex-based comparisons between the two species revealed no significant differences in the condition factors of males, while the condition factors of females were significantly different, suggesting better population health among P. japonicus females compared to P. stimpsoni females. This may be due to the higher number of berried P. stimpsoni at the time of capture. The condition factors of the two spiny lobster species likely reflect the combined effects of zero fishing mortality, ideal rocky habitats along the Jeju Island coast, and nutrient-rich submarine groundwater discharge supporting higher trophic conditions (Kim et al., 2022). Overall, the information gathered on the population status of spiny lobsters can inform sustainable population management efforts.
Molting and molting frequency in decapod crustaceans are regulated by various intrinsic and extrinsic factors (Mykles, 2021). A pair of Y-organs produces molting hormones (ecdysteroids) in response to internal and external stimuli, such as injuries or tissue loss. Molting frequency typically decreases as lobsters increase in size and age (Kulmiye & Mavuti, 2005). Molting plays a key role in spiny lobster fisheries and reproduction, as soft-shelled lobsters are less likely to enter traps and may remain in shallow crevices to avoid predation and cannibalism (MacDiarmid, 1989). In the present study, molting predominantly occurred at night, consistent with previous findings (MacDiarmid, 1989). Two main molting periods were identified: one in May and another from August to October. However, molting frequency decreased with increasing lobster size, potentially explaining the higher molting occurrence in P. stimpsoni, as P. japonicus exhibited larger body sizes in terms of weight and carapace length, particularly among females (Fig. 2). Additionally, the high number of berried P. stimpsoni females may have contributed to the frequent molting, as soft-shelled females are thought to have higher reproductive success (MacDiarmid, 1989). Studies suggest that molting is influenced by colder (Cobb & Phillips, 1980) or warmer (Thomas, 1972) temperatures, and in this study, molting occurred during the warmer months of May to October. Further research is required to investigate the effects of temperature and other external factors on molting behavior.
The study of egg stages focused on P. stimpsoni due to the limited number of berried P. japonicus females in the collected samples. According to Butler IV et al. (2015), since maternal size does not influence egg characteristics, this study did not examine the relationship between egg size and maternal size. The mean diameter at each developmental stage was significantly different, with Stages 1, 2, and 4 showing an increasing trend in egg diameter. However, the mean egg diameter at Stage 3 was significantly smaller than that at Stage 2. This result contradicts the findings of Chang et al. (2007) for Panulirus penicillatus, where egg diameter continuously increased until hatching. Conversely, a study on Panulirus polyphagus reported a decrease in egg diameter with progressive development, a temperature-dependent process (Ikhwanuddin et al., 2014). The reduction in egg diameter at Stage 3 may be attributed to energy consumption and the synthesis of complex cellular and structural compounds by the larvae. However, few studies have been conducted on the egg development of these two species. Therefore, further research is recommended to explore embryonic development and the external and internal factors influencing this process in both lobster species.
The findings of this study are expected to contribute to the sustainable management of these marine resources by providing scientific evidence for implementing fishing quotas and harvest regulations, particularly for berried females, reproductive males, and during molting periods.
Conclusion
This study provides valuable insights into the population distribution, morphometric relationships, condition factors, molting behavior, and egg development stages of Panulirus japonicus and P. stimpsoni in South Korea. Both species show strong correlations between carapace length and body weight, indicating negative allometric growth, with allometric growth coefficients remaining below 3.0. P. japonicus exhibits a more pronounced allometric growth trend than P. stimpsoni. The condition factors consistently exceed 1.00, with female P. japonicus displaying notably higher values than female P. stimpsoni. Molting behavior also differs between the species, with P. stimpsoni primarily molting in May and between August and October, while P. japonicus molts less frequently. Additionally, four distinct stages of egg development were identified in P. stimpsoni, which reaches Stage 4 approximately one month later. These findings help bridge knowledge gaps about these spiny lobsters in South Korea, providing crucial information for their sustainable management and conservation. Further research into the biology, ecology, and population dynamics of these species is recommended to support more effective resource management strategiesies.