






Introduction
Understanding and conserving marine ecosystems is crucial for sustainable fisheries management and marine ecosystem conservation. The coastal waters of Nangdo island, located at the entrance to the semi-enclosed Yeoja Bay in the central South Sea of Korea, are home to a diverse array of fish species (Kim et al., 2015; Lee et al., 2011; Ryu et al., 2011; Yu et al., 2020), playing a crucial role as spawning and nursery grounds during the early life stages of fish (Kim et al., 2015; Moon et al., 2023; Ryu et al., 2011). The composition and community structure of fish in the coastal waters of Yeosu exhibit distinct seasonal variations (Jeong et al., 2005; Kim et al., 2017; Yu et al., 2020), closely linked to changes in marine environmental conditions such as water temperature, salinity, and prey availability (Jeong et al., 2005; Kim et al., 2015; Moon et al., 2022).
Previous studies in the Yeosu coastal waters have shown that the fish species composition and community structure here are characterized by fluctuations in the number of species and individuals present depending on the season. In a previous study focusing on the Nangdo island coastal waters, it was also observed that the fish community structure is regulated by changes in the abundance of migratory species like Engraulis japonicus and Pennahia argentata, as well as resident species like Amblychaeturichthys hexanema and Leiognathus nuchalis (Yu et al., 2020). Furthermore, the Nangdo island coastal area is influenced by its geographical characteristics and seasonal oceanographic factors, with the semi-enclosed Yeoja Bay to the north and the open waters of the Bodol Sea to the south (Cho, 2011). Although Nangdo island, situated at the entrance to Yeoja Bay, is expected to experience various changes in fish species composition and community characteristics depending on the season, surveys were only conducted in May, August, and November of 2017 (Yu et al., 2020). Therefore, elucidating the species composition and community characteristics of fish caught with long-bag stow nets in the Nangdo island coastal waters is expected to contribute to the management and conservation strategies for fish resources in the surveyed area (Froese & Pauly, 2024).
Recently, climate change has led to rising global ocean temperatures (IPCC, 2021), impacting coastal ecosystems like Yeoja Bay. Furthermore, rising temperatures can influence the physiological traits, habitat distribution, and spawning seasons of fish, potentially leading to significant changes in fish community structures (Perry et al., 2005; Pörtner & Peck, 2010). Specifically, there may be an increase in warm-water species, a decrease in cold-water species, changes in spawning seasons, and northward fish migration (Cheung et al., 2009; Lee et al., 2021; Rijnsdorp et al., 2009). Therefore, measures for managing and conserving Yeoja Bay’s fishery resources in response to climate change are essential, necessitating continuous monitoring of fish species composition and community dynamics in this important spawning and nursery ground.
In this study, we utilized catch data from a long-bag stow net located in Nangdo island at the entrance of Yeosu Bay, excluding the fishing temporary closure (January-February) and the closed season (May). First, we analyzed the seasonal species composition of fish caught in the coastal waters of Nangdo island, Yeosu, and identified changes in the fish species appearing in each season. Second, we investigated the seasonal variations in fish community structure and the influence of marine environmental factors (e.g, water temperature and salinity). Third, we analyzed the coastal recruitment characteristics of the robust tonguefish (Cynoglossus robustus), an important commercial species in the South Sea, to understand its distribution and habitat. This study aims to provide baseline data for fish resource management and marine ecosystem conservation in the Nangdo island coastal waters, as well as to obtain valuable information for predicting and responding to future changes in fish community structure due to marine environmental changes.
Materials and Methods
Water temperature and salinity around the Nangdo island long-bag stow net were measured using a CTD diver (vanEssen, Waterloo, Canada) and an RBRconcerto3 (conductivity, temperature, and depth; RBR, Ottawa, Canada) at the location where the long-bag stow net was installed.
This study was conducted once a month from March to December 2021, excluding the fishing temporary closure (January-February) and the closed season (May), using a long-bag stow net installed in the coastal waters of Nangdo island, Hwajeong-myeon, Yeosu-si, Jeollanam-do province, Korea (Fig. 1). The long-bag stow net, a stationary fishing gear with long bag-shaped nets fixed with anchors at the wings and the end of the bag, allowing it to move with the current, had a total length of 25 m, a net opening of 10 m in both width and height, a mesh size of 5 cm, and a cod-end mesh size of 1 cm. The catch was kept in ice boxes to maintain freshness and transported to the laboratory. Fish caught in the long-bag stow net were identified to the species level following Kim et al. (2005a), and the taxonomic system and scientific names followed FishBase (Froese & Pauly, 2024). The total length of each classified fish species was measured to the nearest 0.1 mm, except for hairtail, for which anal length was measured. Body weight was measured to the nearest 1 gram. In this study, only the abundance of captured fish was analyzed, as weight measurements were excluded to emphasize species diversity, community structure, and the recruitment characteristics of the long-bag stow net fishery.
To understand the fish species diversity in the Nangdo island coastal waters, the Shannon-Wiever diversity index (H’) was calculated (Shannon & Weaver, 1963). Pearson correlation analyses were performed using the “vegan” and “corrplot” packages in R v4.2.3 to explore the possible between the C. robustus, temperature, salinity, and three major shrimps (Trachysalambria curvirostris, Batepenaeopsis tenella, and Metapenaeus joyneri). To analyze community characteristics, multivariate analysis techniques, including classification and ordination, were applied to taxa that appeared with a frequency of 5% or more among the total number of individuals caught. The number of individuals of each fish species used in the community analysis was log-transformed (Logx + 1) to normalize the data and avoid bias due to differences in sampling time and population density among species. To measure the monthly similarity of fish communities, hierarchical cluster analysis was performed using the Bray-Curtis similarity index and the unweighted pair groups method with arithmetic mean (UPGMA), and the results were visualized using non-metric multidimensional scaling (nMDS). The species influencing each group identified in the fish community analysis were analyzed using similarity percentages procedure (SIMPER). The similarity between groups was verified using the Analysis of Similarities (ANOSIM) test. All analyses were conducted using PRIMER (version 6) software (Clarke et al., 2014). Additionally, the monthly length composition of C. robustus, a commercially significant species captured using long-bag stow nets at the entrance of Yeoja Bay, was analyzed to understand its recruitment characteristics into the bay. The study also examined the relationship between this species and shrimp, one of its main prey items, to understand how prey availability might influence recruitment patterns.
Results
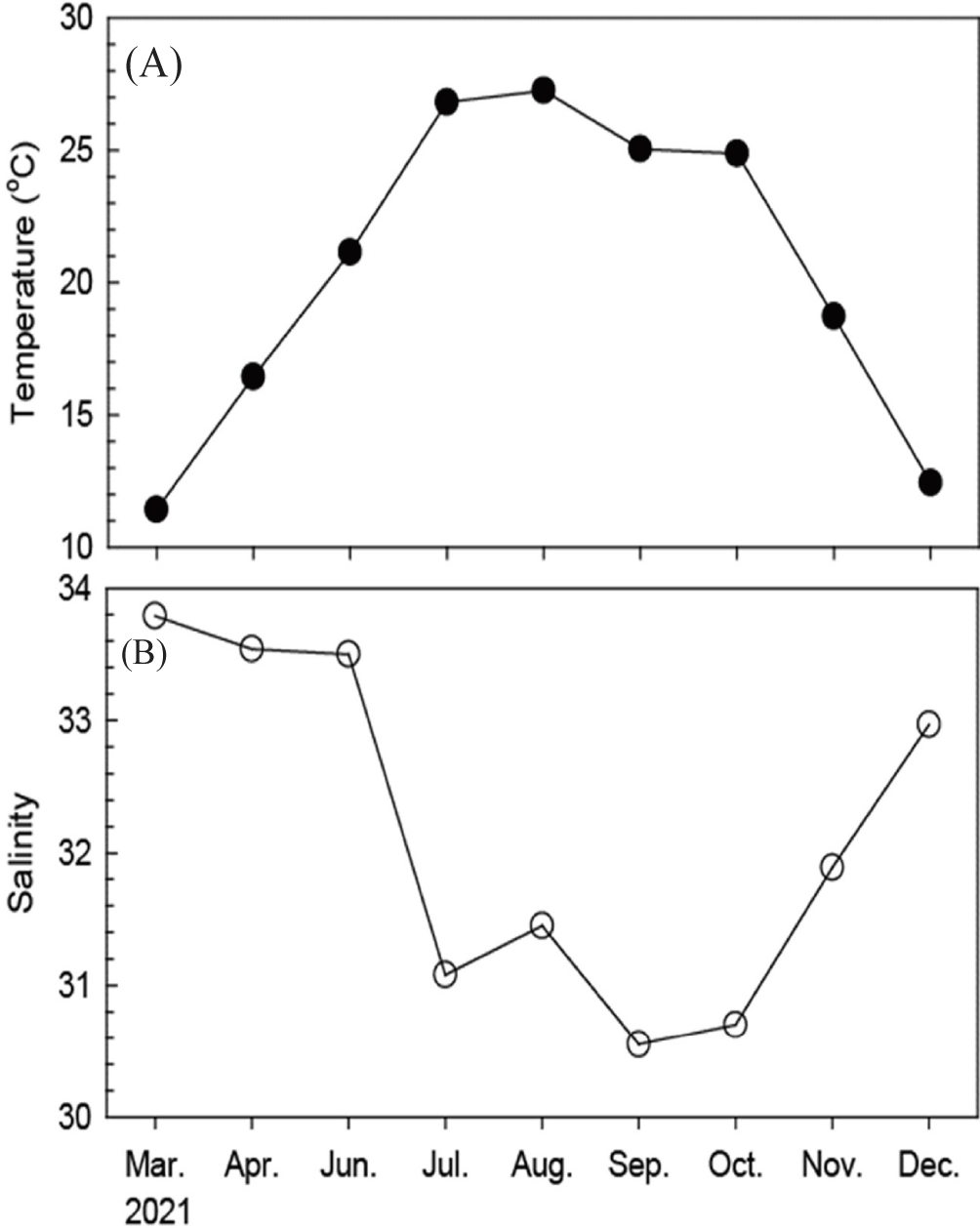
During the survey period, the monthly water temperature in the coastal waters of Nangdo island ranged from 11.4°C to 27.3°C, with the lowest in February (winter) and the highest in August (summer; Fig. 2A). Monthly salinity ranged from 30.5 to 33.8, with the lowest in September (autumn) and the highest in March (spring; Fig. 2B). The fluctuations in water temperature and salinity showed typical seasonal variations.
During the survey period, excluding the fishing temporary closure and the closed season, a total of 169,798 fish individuals were collected, representing 10 orders, 41 families, and 69 species (Table 1). The most abundant order was Perciformes with 19 families and 31 species, followed by Scorpaeniformes (8 families, 8 species), Pleuronectiformes (3 families, 7 species), Clupeiformes (2 families, 7 species), Tetraodontiformes (1 family, 4 species), and others (9 orders, 11 families, 13 species). In terms of the number of individuals, Johnius grypotus was the most dominant species, accounting for 27.5% of the total with 46,750 individuals. The next most dominant species were E. japonicus (38,702 individuals, 18.6%), Thryssa kammalensis (28,905 individuals, 13.9%), Trichiurus japonicus (22,320 individuals, 10.7%), P. argentata (12,250 individuals, 5.9%), L. nuchalis (4,514 individuals, 2.2%), Pampus echinogaster (2,134 individuals, 1.0%), Thryssa hamiltonii (1,727 individuals, 0.8%), C. robustus (1,484 individuals, 0.7%), and the remaining species accounted for less than 0.5% of the total.
The monthly number of species observed ranged from 18 to 37, with the highest number (37) in November (autumn) and the lowest (18) in August (summer). The species composition showed an increasing trend from summer to autumn (Table 1 and Fig. 3A). The monthly number of individuals caught varied greatly, ranging from 675 to 70,876 (Table 1 and Fig. 3B), with the highest in April (spring) and the lowest in December (winter). In March, 26 species were observed, with Acropoma japonicum, T. kammalensis, Setipina tenuifills, and Takifugu niphobles being the most abundant. In April, the highest number of individuals was recorded due to an increase in the recruitment of E. japonicus and T. kammalensis under 8 cm in fork length. In June, the recruitment of Trichiurus japonicus increased, resulting in a high catch, while that of E. japonicus decreased compared to April. The dominant species in June were J. grypotus, T. kammalensis, and Pampus echinogaster. In July, the recruitment of J. grypotus increased, making it the most dominant, followed by T. hamiltonii, Trachurus japonicus, Larimichthys polyactis, and Odontamblyopus lacepedii. In August, the hottest month, the recruitment of J. grypotus further increased, leading to a very high catch. The next dominant species were Pampus echinogaster, O. lacepedii, and Chaeturichthys stigmatias. In September, the recruitment of J. grypotus decreased, while that of P. argentata increased. The next most abundant species were J. grypotus, S. tenuifills, Apogon lineatus, and C. robustus. October showed a similar pattern to September, with J. grypotus, P. argentata, C. robustus, and L. nuchalis being dominant. In November, besides the dominant J. grypotus and P. argentata, the recruitment of Sillago japonica increased. The next dominant species were C. robustus and Thryssa hamiltonii. In December, the number of individuals caught decreased significantly, with Lagocephalus wheeleri being the most dominant, followed by Dasyatis akajei, O. lacepedii, and Pagrus major.
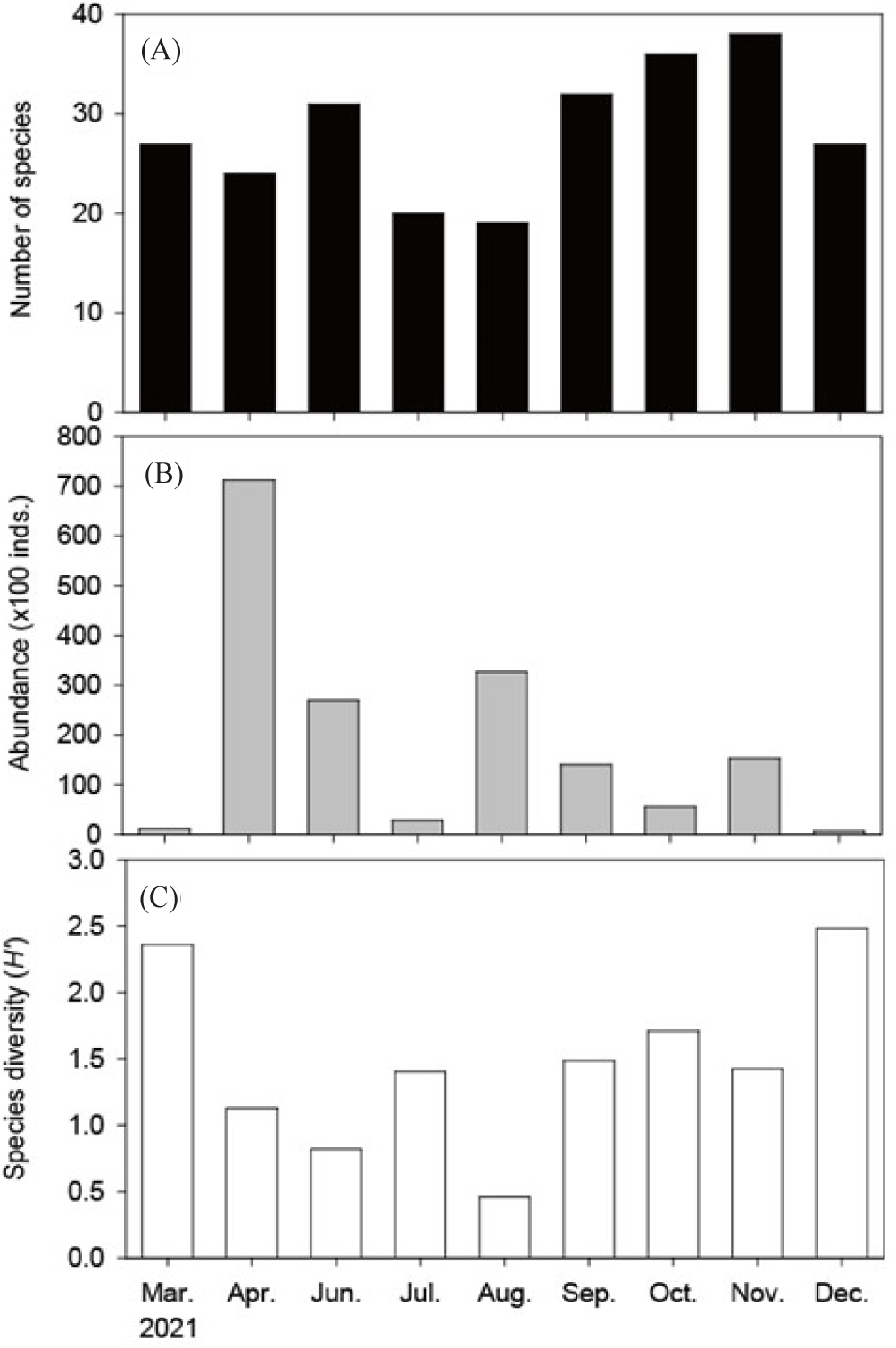
The monthly Shannon diversity index (H’) ranged from 0.45 to 2.48. Despite having 18 species present in August, the diversity index was the lowest. The highest diversity index was observed in December, which also had the highest number of species observed (Fig. 3C).
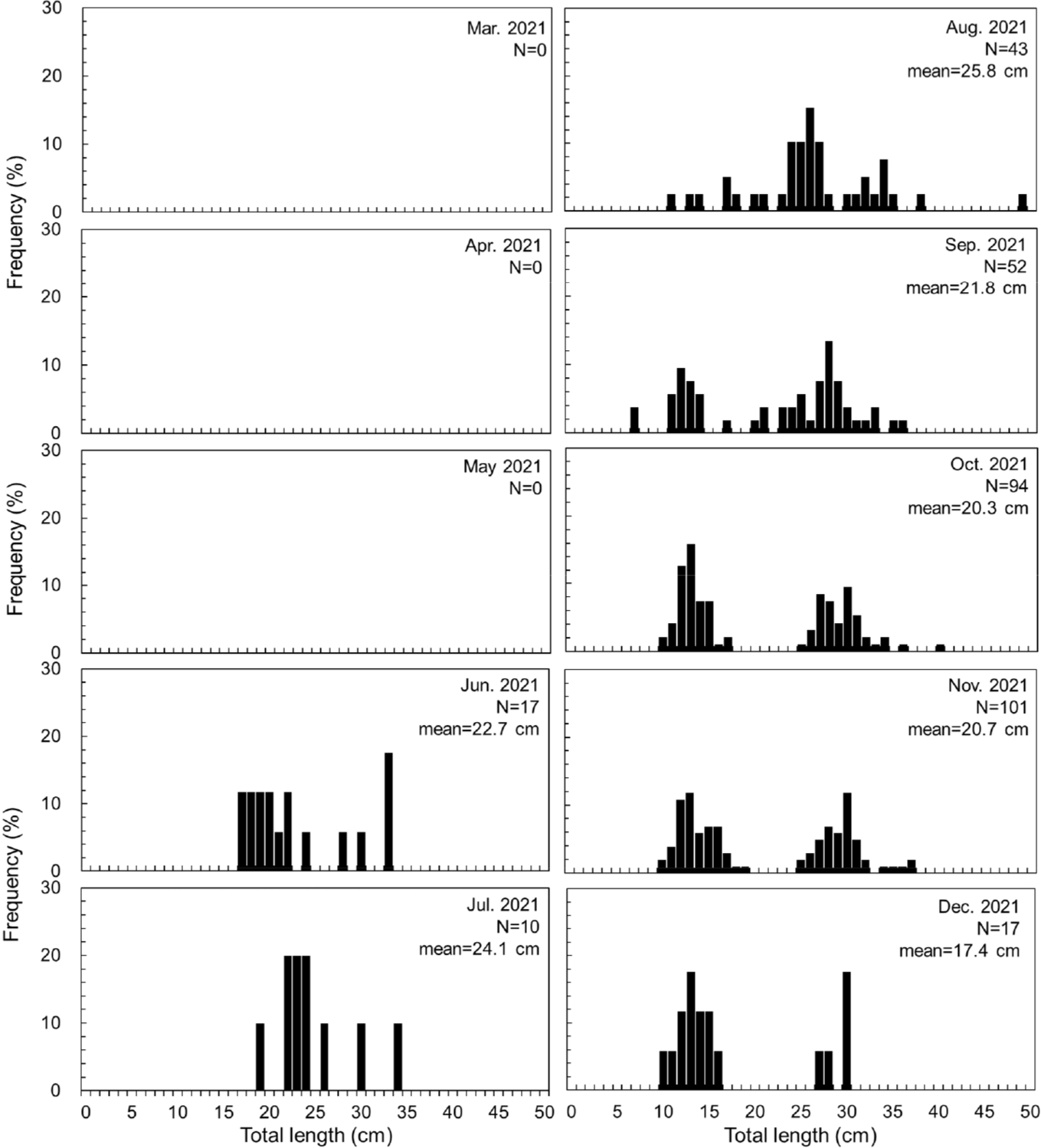
During the survey period, C. robustus, a commercially important fish species, continuously migrated into the Yeoja Bay from June, except for the fishing temporary closure and the closed season. The monthly total length (TL) distribution of the fish showed a range of 6.8–48.1 cm (Fig. 4). The monthly TL mode showed a mode in the range of 17–22 cm in June, and a clear mode above 20 cm during the spawning season of C. robustus (July-August). From September to December, two modes were continuously observed in the 10–15 cm and 25–30 cm ranges as the recruitment into the Yeoja Bay increased. In particular, from September, after the spawning season (June-August), the proportion of small individuals under 15 cm increased, and they continued to migrate into the Yeoja Bay until December.
An investigation into the appearance patterns of shrimp species, which serve as key prey for C. robustus (Fig. 5A), and their recruitment into Yeoja Bay revealed that the abundance of Trachysalambria curvirostris (Fig. 5B), B. tenella (Fig. 5C), and M. joyneri (Fig. 5D) was high from June to November, coinciding with the period when C. robustus entered the bay. Notably, the peak abundance of these three shrimp species was observed in September, which also marked the period when Cynoglossus joyneri individuals with a TL under 15 cm were present. Analysis of the abundance of total shrimps and M. joyneri indicates that C. robustus exhibits a broader tolerance range for both total shrimps and M. joyneri (Fig. 5D).
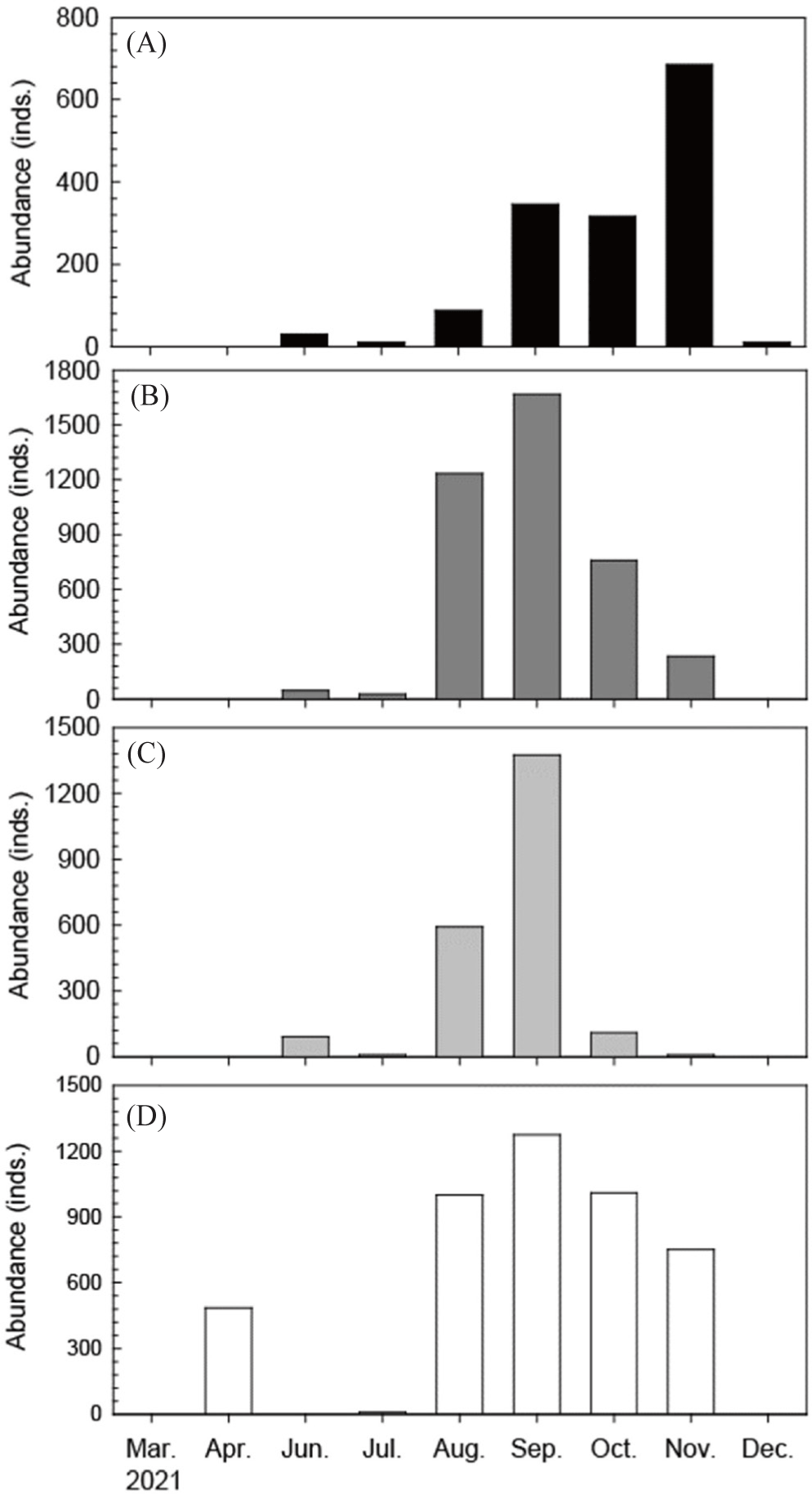
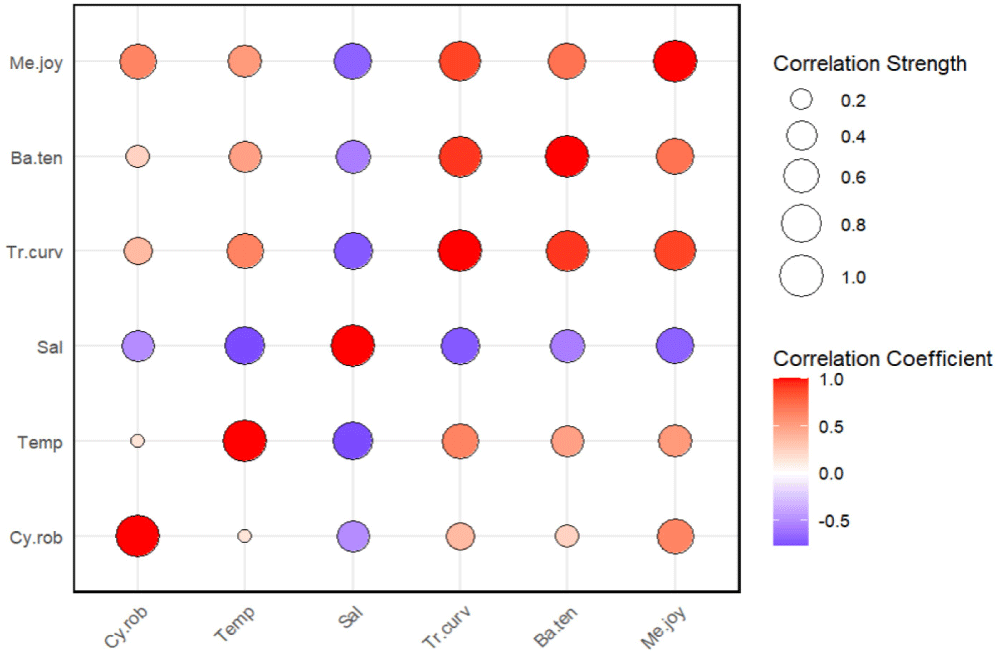
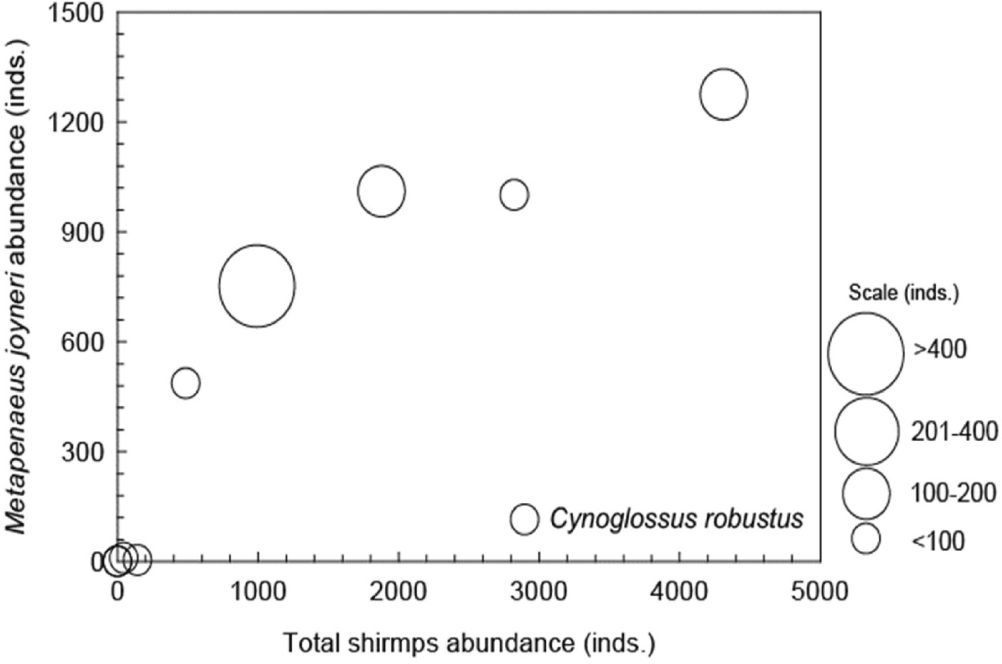
A Pearson correlation analysis was conducted to examine the relationships between C. robustus and environmental factors, including temperature, salinity, and three major shrimp species (Trachysalambria curvirostris, B. tenella, and M. joyneri). The analysis revealed that C. robustus had the strongest positive correlation with M. joyneri, showing a correlation coefficient of 0.621. In contrast, salinity exhibited a moderate negative correlation with C. robustus, while the remaining factors displayed weak or negligible correlations (Fig. 6). Additionally, C. robustus demonstrates a more restricted occurrence pattern, with higher abundances observed in association with elevated total shrimp levels. Moreover, C. robustus shows a preference for M. joyneri abundance ranging from 600 to 1,300 individuals (Fig. 7).
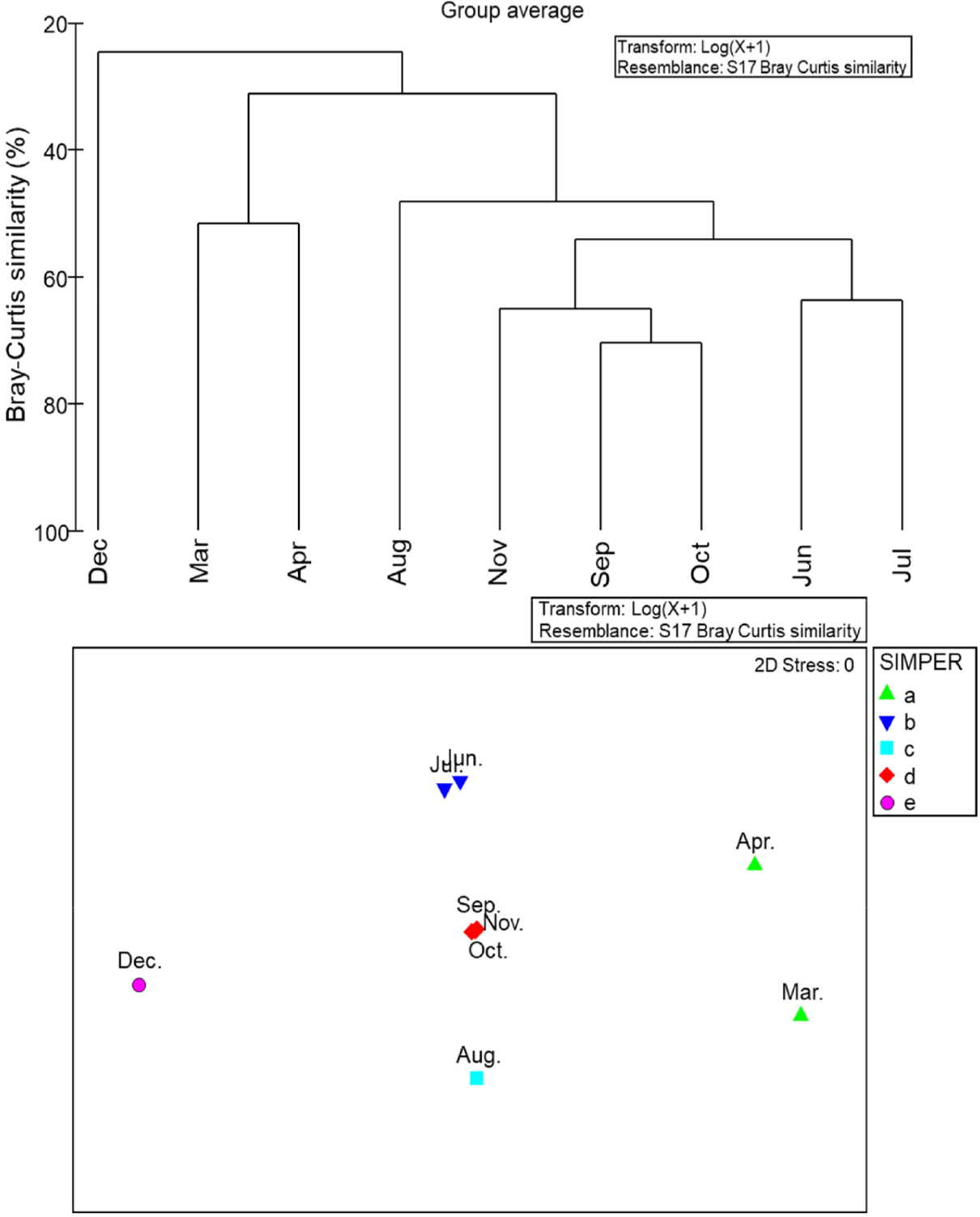
Based on the number of individuals caught in the long-bag stow net located near Nangdo island at the entrance of Yeoja Bay, cluster analysis and nMDS ordination revealed six distinct clusters with a Bray-Curtis similarity index of 50% (Fig. 8). The taxa that influenced each cluster were as follows: Cluster A (March-April): A. japonicum, T. kammalensis, S. tenuifills, Takifugu niphobles, Cluster B (June-July): J. grypotus, Thryssa hamiltonii, Trichiurus japonicus, O. lacepedii, Cluster C (August): J. grypotus, Pampus echinogaster, O. lacepedii, Cluster D (September-November): J. grypotus, P. argentata, C. robustus, Thryssa hamiltonii, Cluster E (December): Lagocephalus wheeleri, D. akajei, O. lacepedii, Pagrus major (Table 2). ANOSIM analysis to assess the similarity between clusters showed significant differences between groups at a 95% confidence level (Global test 0.935, p < 0.05).
Discussion
This study aimed to provide baseline data for fisheries resource management and marine ecosystem conservation by investigating the community structure of fishes and recruitment characteristics of the commercially important tonguefish (C. robustus) in long-bag stow net catches near Nangdo Island, located at the entrance of Yeoja Bay. During the survey period, a total of 10 orders, 41 families, and 69 species were observed. Yeoja Bay exhibited typical seasonal variations, with low water temperatures and high salinity in winter shifting to high water temperatures and low salinity in summer (Moon et al., 2023; Park et al., 2011). These seasonal changes in water temperature and salinity influence the physiological characteristics, habitat selection, and prey distribution of fish, thus acting as major environmental factors driving seasonal changes in fish community structure (Cheung et al., 2009; Moon et al., 2024). The species composition and community of fish entering Yeoja Bay were dominated by small forage fish such as A. japonicum and T. kammalensis in spring, and by demersal fish such as J. grypotus, Pampus echinogaster, Larimichthys polyactis, O. lacepedii, and C. robustus from summer to autumn, indicating distinct seasonal community changes. These seasonal community changes reflect broader ecological dynamics and underscore the influence of environmental conditions on fish life cycles.
Seasonal changes in fish communities have significant implications beyond species composition, affecting various aspects such as fish life history, food web structure, and ecosystem function (Blaber, 2000). In spring, as water temperatures rise after winter, spring-spawning fish become more active, migrating to the southern coast and inner bays for spawning (Kim et al., 2007; Moon et al., 2022). Consequently, the density of zooplankton, an important food source for spring-spawning fish, increases in Yeoja Bay (Moon et al., 2023). During this period, small pelagic fish such as E. japonicus, T. kammalensis, and Setipinna tenuifilis, which primarily feed on zooplankton are dominant. In contrast, as temperatures rise in summer, the number of species observed decreases, while the abundance of demersal fish such as Larimichthys polyactis, Trichiurus japonicus, and Thryssa hamiltonii increases. Notably, J. grypotus showed a marked dominance during period from April to July, and recruitment of Larimichthys polyactis also increased. This is likely because these species, whose spawning season occurs during this period, migrate to the coast and inner bays for spawning (Kang et al., 2020; Lee & Park, 1992). In autumn, as temperatures gradually decrease, fish activity lessens, and feeding increases to store energy for winter or spawning (Gibson, 1994; Reeve et al., 2022). During this time, demersal fish that feed on benthic organisms become dominant (Gibson, 1994; Ziegler et al., 2019). The recruitment of P. argentata with a total length of 2.6–23.6 cm increased from September to December, suggesting that P. argentata, a demersal fish, migrates to Yeoja Bay for spawning from May to July (Jeon et al., 2020). Both parents and juveniles appear simultaneously even after spawning, indicating that Yeoja Bay serves as a spawning and nursery ground for P. argentata. Conversely, Odontamblyopus lacepedii, which appears monthly at the entrance of Yeoja Bay during the survey period, has a spawning season from June to July (Park et al., 2007). The number of individuals of this species increased from post-spawning season to August, likely due to increased spawning individuals in Yeoja Bay, where the well-developed tidal flats provide an ideal habitat for Odontamblyopus lacepedii, which burrows and resides in such environments.
In this study, C. robustus continuously recruitment into Yeoja Bay except for January to May, and notably showed an increase in the recruitment of small individuals after the spawning season. Monthly TL data showed that individuals larger than 20 cm entered during the spawning season (July to August; Baeck & Kim, 2004), with the proportion of small individuals under TL 15 cm increasing from September and continuing until December. Yeoja Bay includes diverse habitats such as tidal flats and seagrass beds, which offer abundant food and shelter for young fish (Kim et al., 2015; Moon et al., 2023). The plentiful zooplankton (Lomartire et al., 2021; Moon et al., 2023), polychaetes (Kim et al., 2005b), and shrimps (Kim et al., 2007), which are major food sources for these fish (Baeck & Huh, 2004), likely support the growth of C. robustus entering Yeoja Bay. This suggests that Yeoja Bay serves as a critical spawning and nursery ground for C. robustus, a commercially important species in the southern coast of Korea (Moon et al., 2022). Thus, to ensure the sustainable use of C. robustus resources, effective management measures are essential. These should include restricting the catch of small individuals and adjusting fishing practices during the spawning season (Hilborn & Walters, 1992). Additionally, conserving and enhancing the Yeoja Bay environment is crucial for managing the habitat and nursery grounds of C. robustus.
This study provides valuable baseline data for understanding the fish community structure and recruitment characteristics of the commercially important C. robustus in long-bag stow net catches near Nangdo Island, located at the entrance of Yeoja Bay. The results confirm that Yeoja Bay functions as both a habitat and a nursery ground for various fish species depending on the season, with a particularly crucial role as a nursery ground for C. robustus. However, factors such as rising water temperatures due to climate change, the decline of seagrass beds that serve as nursery grounds, and overfishing of spawning individuals and juvenile fish can impact the Yeoja Bay ecosystem and fish resources. Therefore, to conserve the biodiversity of Yeoja Bay and ensure the sustainable use of fish resources, the following measures are necessary: first, long-term monitoring of changes in fish communities and stock fluctuations of C. robustus; second, conservation of tidal flats and seagrass beds in Yeoja Bay, which provide essential habitats and nursery grounds for fish; third, prevention of overfishing of spawning individuals and juveniles by restricting the catch of small individuals and adjusting fishing efforts during the spawning season. Although this study offers important baseline data for understanding the fish community structure and recruitment characteristics of C. robustus in the waters near Nangdo Island, only limited environmental factors such as water temperature and salinity were considered. Future research should explore the effects of various environmental factors, including dissolved oxygen, nutrients, zooplankton, and seagrass beds on fish communities, and conduct additional surveys to investigate C. robustus eggs and larvae in spawning grounds.
Conclusions
This study provides essential baseline data on the community structure and recruitment characteristics of the commercially important tonguefish (C. robustus) in Yeoja Bay, Korea. Seasonal changes significantly influence fish community dynamics, with a noticeable transition from small pelagic fish in spring to demersal fish in summer and autumn. Yeoja Bay serves as an essential spawning and nursery ground for C. robustus, evidenced by marked increase in the recruitment of small individuals following the spawning season. The diverse habitats in Yeoja Bay, such as tidal flats and seagrass beds, play a vital role in supporting the growth and survival of juvenile fish. However, rising water temperatures and habitat degradation driven by climate change pose serious threats to this ecosystem. To ensure the sustainability of C. robustus populations and preserve the biodiversity of Yeoja Bay, it is imperative to implement long-term monitoring programs, conserve key habitats, and enforce stricter regulations on fishing activities, particularly during spawning seasons. Future research should investigate additional environmental factors affecting fish communities and focus on the early stages of C. robustus to enhance management strategies.