






Introduction
The family Pinguipedidae includes 7 genera and 101 species globally (Fricke et al., 2025), with 3 genera and 30 species in Japan (Motomura, 2020), and 1 genus with 6 species in Korea (Marine Biodiversity Institute of Korea [MABIK], 2023; Park et al., 2007). The genus Parapercis is the largest within the Pinguipedidae, including approximately 54 species worldwide (Froese & Pauly, 2024; Randall & Francis, 1993). The pigmentation or coloration patterns serve as one of the most useful diagnostic keys for distinguishing between species within this genus (Sparks et al., 2021). These species are distributed globally, with 51 species in the Indo-West Pacific and individual species in the Southeast Pacific, South Atlantic, and Northeast Atlantic (Allen & Erdmann, 2017; Cantwell, 1964; Johnson, 2006; Randall & McCosker, 2002; Randall, 2003; Randall & Yamakawa, 2006). They inhabit rocky or sandy subtidal areas at depths of about 100 m and primarily feed on invertebrate benthic macroinvertebrates (Kim et al., 2005; Randall et al., 2008; Yamada et al., 2007); they lay pelagic eggs (Allen & Erdmann, 2017; Froese & Pauly, 2024) and some of them exhibit protogynous hermaphroditism (Marshall, 1950; Nakazono et al., 1985; Randall et al., 2008; Stroud, 1984).
According to MABIK (2023) and the NIBR (2023), 6 Parapercis species are officially recorded in Korea: Parapercis aurantiaca, Parapercis multifasciata, Parapercis muronis, Parapercis pulchella, Parapercis sexfasciata, and Parapercis snyderi. On February 27, 2024, a specimen of Pinguipedidae was collected off the southwestern coast of Seogwipo-si, Jeju-do Island, Korea, via Danish seine. It was morphologically identified as Parapercis decemfasciata (Franz, 1910), which was confirmed by comparing the mtDNA cytochrome c oxidase subunit I (COI) nucleotide sequence. Although Kim et al. (2005) briefly recorded P. decemfasciata for the first time in their illustrated book, MABIK (2023) and NIBR (2023) did not accept this record due to the absence of voucher specimens. Consequently, we believe our finding is to be a reliable record of P. decemfasciata in Korea, and propose the adoption of the Korean name “No-rang-yeol-dong-ga-ri” as suggested by Kim et al. (2005).
Materials and Methods
The single specimen of Pinguipedidae used in this study was collected using a Danish seine on February 27, 2024, at 33°10’ N, 125°55’ E, off the southwestern coast of Seogwipo-si, Jeju-do Island, Korea (Fig. 1). Collected individual was transported to the laboratory, morphologically identified using Kazuhiko (2013), and after photography, initially fixed in 15% formalin, then washed in running water for 24 h, and finally embedded in 70% ethyl alcohol (EtOH) before being registered in the Ichthyology Laboratory collection of Pukyong National University (PKU) and after deposited in the MABIK.
Terminology and morphometric characteristics for each part of the specimen followed the methods of Cantwell (1964) and Hubbs & Lagler (2004), and eight meristic characters, including dorsal fin rays, and 19 morphometric characters, including total length (TL), were analyzed. Morphometric characters were measured to the nearest 0.01 mm using Vernier calipers. The measurements for each body were expressed as a percentage (%) of the standard length (SL) and head length (HL). In addition, we additionally counted, measured, and tabulated Parapercis sexfasciata (PKU 54187), and Parapercis multifasciata (PKU 57583) for morphological comparisons with P. decemfasciata (Table 1).
| Species | Parapercis decemfasciata | Parapercis sexfasciata | Parapercis multifasciata | Parapercis aurantiaca |
|---|---|---|---|---|
| MABIK PI00062187 (Present study) | PKU 54187 (Present study) | PKU 57583 (Present study) | Youn (1998) | |
| Total length (mm) | 128.67 | 140.06 | 132.71 | - |
| Standard length | 108.12 | 120.54 | 113.35 | 127.95 |
| Measurements | ||||
| As % of standard length | ||||
| Head length | 26.74 (24.73) | 31.69 (26.29) | 30.53 | 25.60 |
| Interorbital width | 3.17 (2.93) | 4.40 (3.65) | 4.06 | - |
| Postorbital length | 11.02 (10.19) | 11.25 (9.33) | 7.84 | - |
| Dorsal fin base length | 67.15 (62.11) | 75.45 (62.59) | 66.24 | - |
| Preanal length | 45.87 (42.43) | 51.18 (42.46) | 49.78 | 47.80 |
| Predorsal length | 31.40 (29.04) | 34. 09 (28.28) | 35.33 | 30.90 |
| Prepelvic length | 24.35 (22.52) | 27.14 (22.52) | 30.43 | - |
| Anal fin base length | 51.31 (47.46) | 54.88 (45.53) | 46.88 | - |
| As % of head length | ||||
| Snout length | 6.59 (24.64) | 8.99 (28.37) | 10.08 | 31.50 |
| Orbit length | 8.37 (31.30) | 8.21 (25.91) | 8.42 | 36.30 |
| Upper jaw length | 9.87 (36.91) | 11.09 (35.00) | 12.05 | - |
| Body depth | 18.22 (68.14) | 21.54 | 20.70 | 20.80 |
| Body width | 15.84 (59.24) | 14.62 | 14.83 | - |
| Caudal peduncle depth | 9.89 (36.99) | 9.93 | 10.82 | - |
| Longest pectoral fin length | 20.91 (78.20) | - | 22.53 | 76.90 |
| Longest pelvic fin length | 17.75 (66.38) | 19.75 | 21.61 | 74.20 |
| Longest anal fin length | 11.25 (42.07) | 16.26 | 18.90 | 45.00 |
| Counts | ||||
| Dorsal fin | IV, 23 | V, 23 | V, 23 | V, 23 |
| Pectoral fin | 19 | - | 19 | 20 |
| Pelvic fin | I, 5 | I, 5 | I, 5 | - |
| Anal fin | I, 19 | I, 19 | I, 19 | I, 19 |
| Lateral line scales | 57 | 62 | 60 | 64 |
| Lateral line pores | 52 | 62 | 53 | - |
| Vertical bands on caudal fin | 4 | - | 7 | - |
| Oblique bands | 10 | 6 | 10 | - |
The values in parentheses represent the proportions according to standard length (SL) and head length (HL), respectively.
Data from Hubbs & Lagler (2004), Kazuhiko (2013).
Muscle tissues of one specimen of Pinguipedidae used in this study were dissected and stored in 99% ethyl alcohol, and genomic DNA was extracted using Chelex 100 resin (Bio-rad, Hercules, CA, USA). Amplification of the mitochondrial DNA cytochrome oxidase subunit I (mtDNA COI) region was performed using FishF1 (5’-TCA ACC AAC CAC AAA GAC ATT GGC AC-3’) and FishR1 (5’-TAG ACT TCT GGG TGG CCA AAG AAT CA-3’) primers (Ward et al., 2005). PCR amplification was performed by polymerase chain reaction (PCR) using a Thermal Cycler (T100™; Bio-rad) under the following conditions: [initial denaturation at 95 for 5 min; followed by 35 cycles of denaturation at 94°C for 30 s, annealing at 52°C for 45 s, extension at 72°C for 45 s, final extension at 72°C for 7 min and infinite hold at 4°C]. Mitochondrial DNA COI sequences after the reaction were aligned using ClustalW multiple alignments (Thompson et al., 1994) in BioEdit version 7.2.5 (Hall, 1999). The COI sequence of one P. decemfasciata used in this study has been deposited in the National Center for Biotechnology Information (NCBI) and assigned accession number (MABIK PI00062187: PQ277024). The genetic tree was calculated with the Kimura 2-parameter model (Kimura, 1980) using the program MEGA 11.0.13 (Tamura et al., 2021), and a neighbor-joining tree was constructed after 1,000 bootstrap replications. To determine the degree of close relationships between species, we used the NCBI-registered P. decemfasciata (KU944748), Parapercis multifasciata (KU944741), Parapercis muronis (HM422380), Parapercis sexfasciata (LC201782), and Parapercis pulchella (KU944746), and as an outgroup, Pholis crassispina (NC004410).
Results
Neopercis decemfasciataFranz, 1910: 81 (Type locality: Misaki, Japan); Kamohara, 1952: 87 (Japan).

| Species | P. decemfasciata | |||
|---|---|---|---|---|
| MABIK PI00062187 (Present study) | Franz (1910) | Yamada et al. (2007) | Kazuhiko (2013) | |
| Total length (mm) | 128.67 | 90.00–130.00 | 98.00–148.00 | - |
| Standard length | 108.12 | - | 81.00–122.00 | 150.00 |
| Measurements | ||||
| As % of standard length | ||||
| Head length | 26.74 (24.73) | 23.81–25.00 | 24.39–26.32 | - |
| Interorbital width | 3.17 (2.93) | - | - | - |
| Postorbital length | 11.02 (10.19) | - | - | - |
| Dorsal fin base length | 67.15 (62.11) | - | - | - |
| Preanal length | 45.87 (42.43) | - | 38.46–43.48 | - |
| Predorsal length | 31.40 (29.04) | - | - | - |
| Prepelvic length | 24.35 (22.52) | - | - | - |
| Anal fin base length | 51.31 (47.46) | - | - | - |
| As % of head length | ||||
| Snout length | 6.59 (24.64) | - | 21.28–27.78 | - |
| Orbit length | 8.37 (31.30) | 33.33 | 33.33–38.46 | - |
| Upper jaw length | 9.87 (36.91) | - | 37.04–43.48 | - |
| Body depth | 18.22 (68.14) | - | 66.67–83.33 | - |
| Body width | 15.84 (59.24) | - | 62.50–71.43 | - |
| Caudal peduncle depth | 9.89 (36.99) | - | 34.48–40.00 | - |
| Longest pectoral fin length | 20.91 (78.20) | - | - | - |
| Longest pelvic fin length | 17.75 (66.38) | - | - | - |
| Longest anal fin length | 11.25 (42.07) | 45.45 | - | - |
| Counts | ||||
| Dorsal fin | IV, 23 | IV, 22–23 | IV, 23–24 | IV, 23–25 |
| Pectoral fin | 19 | - | 19–20 | 20 |
| Pelvic fin | I, 5 | - | I, 5 | - |
| Anal fin | I, 19 | ?, 19–20 | I, 18–20 | I, 19–20 |
| Lateral line scales | 57 | 57 | 57–63 | - |
| Lateral line pores | 52 | 52 | - | 61 |
| Vertical bands on caudal fin | 4 | 4–5 | 4–6 | - |
| Oblique bands | 10 | - | 10–11 | 10 |
The values in parentheses represent the proportions according to standard length (SL) and head length (HL), respectively.
Data from Hubbs & Lagler (2004), Kazuhiko (2013).
P. decemfasciata: Tomiyama & Abe, 1958: 119 (Japan); Cantwell, 1964: 251 (Japan); Masuda et al., 1984: 291 (Japan); Shen et al., 1993: 488 (Taiwan); Kim et al., 2005: 398 (Japan, Taiwan); Neumann, 2006: 279 (Japan); Yamada et al., 1985: 296 (Japan); Yamada et al., 2007: 895 (Japan); Kazuhiko, 2013: 1257 (East China Sea).
MABIK PI00062187 (Previously: PKU 63100), one individual, 108.12 mm SL, off the southwestern coast of Jeju-do Island, Korea (33°10′41.9″N 125°55′15.0″E), 27 February, 2024, by Danish seine, about 104–106 m depth, collector Do Hyun Oh.
Counts and measurements are shown in Tables 1 and 2.
Body fusiform and elongated. Body covered with ctenoid scales except interorbital region and nasal pores. Head large, pointed with short snout. Eye relatively large, rounded, located on dorsal part of head and slightly longer than snout; interorbital space narrow. Mouth moderately large and terminal, or upper lip slightly protruding beyond tip of lower jaw; oblique with slightly projecting lower jaw; maxilla extending beyond a vertical anterior margin of eye; palatine teeth present with 2 rows. Gill rakers short and rounded. Opercle with a single strong retrorse spine dorsally. Outer margin of preopercle smooth and that of subopercle with faint serrations in the middle portion. Lateral line continuous, extending from opercle to pectoral fin curving slightly upward, and following straight through middle to center of caudal peduncle. Origin of dorsal fin above fourth lateral line scale. Dorsal spines connected by membrane to its base, 4th spine and 14th soft ray of dorsal-fin each longest; Spines and soft rays of dorsal-fin smoothly connected without notch; origin of dorsal fin base posterior to that of pectoral and anal fin base; dorsal fin spine thin and significantly shorter than soft rays. Base of dorsal and anal fin long. Pectoral fin relatively rounded when spread, and its tips extending slightly beyond anus; 10th soft ray longest. Beginning of last pelvic and dorsal-fin ray bifurcated. Appressed pelvic fin extending posterior of anus. Origin of anal fin below base 6th soft ray of dorsal fin, anal-fin spine slender, closely attached to first soft ray; 16th soft ray longest. Caudal fin rounded.
Body bright red overall. Head with black spots scattered throughout. 10 narrow yellowish-brown transverse bands from base of dorsal fin to bottom of lateral line on body side. 4 narrow dark transverse bands on caudal fin; Upper lobe of caudal fin speckled with rounded pale black spots. Each fin yellow except for pectoral fin. (Fig. 2A).
Body light gray overall. Pectoral fin base white. 10 horizontal bands on body side black above lateral line but faint below. Each fin pale gray (Fig. 2B).
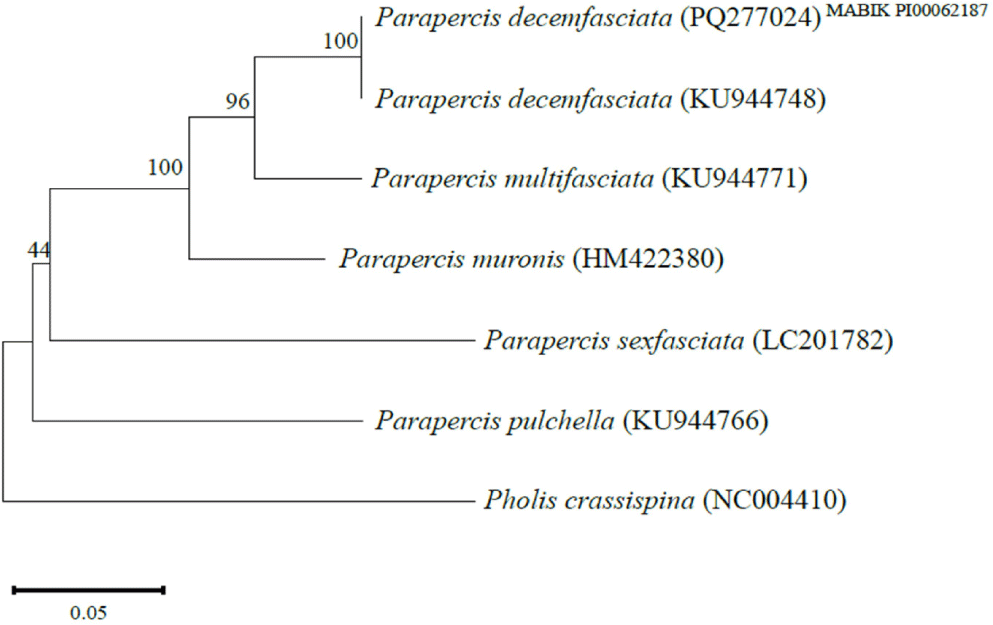
A 529 bp segment of the COI sequence (PQ277024) from the specimen was compared to the genera Parapercis and Pholis registered at the NCBI. The specimen closely matched P. decemfasciata (KU944748) with 100% identity. Parapercis multifasciata (KU944771) clustered with a genetic distance of 0.07, while Parapercis muronis (HM422380), Parapercis sexfasciata (LC201782), and Parapercis pulchella (KU944766) clustered with 0.10, 0.22, and 0.25, respectively. Pholis crassispina (NC004410), used as an outgroup, showed clear divergence from these species with a genetic distance of 0.28 (Fig. 3).
P. decemfasciata occurs in the coastal waters of Jeju-do Island, Korea (This study), off Southern Japan (Kim et al., 2005; Masuda et al., 1984; Yamada et al., 1985, 2007), East China Sea (Kazuhiko, 2013) and Taiwan (Kim et al., 2005; Shen et al., 1993).
Discussion
A single specimen of P. decemfasciata was collected off the southwestern coast of Seogwipo-si Jeju-do Island, Korea, on February 27, 2024. The specimen exhibited external morphological traits of P. decemfasciata, including 4 dorsal fin spines, absence of a notch between the spines and soft rays of the dorsal fin, 10 yellow transverse bands on the body side, and 4 dark transverse bands on the caudal fin (Fig. 2 and Table 1). Molecular analysis of the mtDNA COI sequence region confirmed a 100% match with those of P. decemfasciata from NCBI (Fig. 3).
Neumann (2006) noted that 3 specimens of P. decemfasciata from Aburatsubo Bay, Japan in 1904 and 6 specimens from Sagami Bay, Japan, were initially identified as new species by Franz, designated as syntypes, and first described. Unfortunately, these specimens were destroyed during World War II in 1944, leading to sparse information on P. decemfasciata. Very few studies have focused on its taxonomy or have conducted reviews, with only simple checklists or distribution records noted (Cantwell, 1964; Kamohara, 1952; Shen et al., 1993; Shinohara et al., 2001; Yamada et al., 1985, 2007). However, the morphological and genetic characteristics of the specimen collected in this study align well with the defining features of P. decemfasciata, as originally described by Franz in 1910 (Tables 1 and 2).
P. decemfasciata is distinguishable from the closely related species Parapercis multifasciata, based on several morphological and genetic characteristics. The differences include the number of dorsal fin spines (4 in P. decemfasciata vs. 5 in Parapercis multifasciata), the pattern of transverse bands on the body side when fresh (10 light brown bands covering two-thirds of the body side vs. 10 dark brown bands extending to the center of the body side and 10 yellow bands ventrally), and the size relationship between the snout and the pupil (pupil larger than the snout vs. snout larger than the pupil). Additionally, P. decemfasicata lacks the black spot at the base of the caudal fin (present in Parapercis multifasciata) and exhibits no distinctive head markings in contrast to the yellow (H-shaped markings behind the eyes seen in Parapercis multifasciata) (Kim et al., 2005; Kazuhiko, 2013; Yamada et al., 1985, 2007; this study). Furthermore, Cantwell (1964) noted that P. decemfasciata and Parapercis multifasciata, were once considered the same species. However, they show significant differences in the hemal spines, centrum, morphology of the transverse bands in dorsal view, and the groove shape of ctenoid scales before the dorsal fin origin (Yamada et al., 1985, 2007). Additionally, molecular analysis in this study revealed a 7% genetic divergence between the two species, supporting their classification as separate species (Fig. 3). Despite these distinctions, there are very few studies on the description, taxonomic position, and review of P. decemfasciata globally, including Korea. Therefore, further taxonomic research on the Pinguipedidae in Korea is warranted. P. decemfasciata also differs from the congeneric species Parapercis aurantiaca and Parapercis sexfasciata in several aspects: the number of dorsal fin spines (4 in P. decemfasciata vs. 5–6 in both Parapercis aurantiaca and Parapercis sexfasciata), the number of transverse bands on the caudal fin (4 vs. 6 vs. 5), and the number of body side transverse bands when fresh (10 vs. 5 vs. 5–6) (Kim et al., 2005; Shen et al., 1993; Shao, 2024; Youn, 1998; this study).
Conclusion
Since Kim et al. (2005) briefly recorded P. decemfasciata, there have been no official records, leading to uncertainty about its presence in Korean waters. MABIK and NIBR have not included this species in the Korean fish fauna due to the absence of voucher specimens. Based on morphological features such as 4 dorsal fin spines and 10 narrow transverse bands, and the results of molecular identification of our findings, we believe this represents a reliable record of P. decemfasciata from Korea. Consequently, we propose adopting the Korean name “No-rang-yeol-dong-ga-ri” as suggested by Kim et al. (2005).