






Introduction
The genus Labeobarbus (Cyprinidae) is widespread across African freshwater systems, including numerous Ethiopian lakes (Cambray, 1983; Tesfaye & Wolff, 2014). Among its members, the African big barb, Labeobarbus intermedius (Rüppell, 1836), occupies a wide range of habitats, from highland and rift-valley lakes to the Baro–Akobo, Genale–Wabeshebele, and Abay basins (Admassu & Dadebo, 1997; Tesfaye & Wolff, 2014). Notably, Lake Tana, in Ethiopia, supports an endemic species flock of large Labeobarbus, which distinguishes it from other highland lakes (Awoke et al., 2015; de Graaf et al., 2004). Outside of Tana, L. intermedius is common in rift-valley lakes such as Ziway and Hawassa, as well as other Ethiopian freshwater bodies (Tesfaye & Wolff, 2014). Its presence in diverse ecosystems underscores its ecological resilience and significance. Economically, L. intermedius contributes notably to local fisheries, accounting for approximately 9% of Ethiopia’s annual catch (Tesfaye & Wolff, 2014).
L. intermedius from different Ethiopian lakes showed omnivorous feeding habits, mainly feeding on gastropods, phytoplankton, macrophytes, insects, detritus, and nematodes with the tendency to shift to piscivorey (feeding on fish) as it grows older (Admassu & Dadebo, 1997; Desta et al., 2006; Sibbing, 1998). The findings of Kassahun (2005) and Dadebo et al. (2013) from Lake Koka oppose the piscivores feeding habits of the fish, stating that the gut contents were composed of macrophytes detritus, insects, and nematodes.
A substantial body of information exists in regard to L. intermedius feeding behavior in various aquatic habitats of Ethiopia, such as Tana, Hawassa, and Koka lakes. Nevertheless, feeding ecology in riverine habitats of southwestern Ethiopia, especially concentrating on the Beles River, has not been sufficiently investigated. This ignorance is of paramount significance due to the fact that the Beles River is a vital tributary that feeds into the Blue Nile, with direct implications in regard to affecting the Grand Ethiopian Renaissance Dam (GERD) reservoir’s ecological soundness. In addition, riverine habitats are also increasingly being subjected to anthropogenic pressures that could impact food web associations and pose negative influences on key fisheries such as L. intermedius. In this regard, recognizing the trophic ecology of L. intermedius is not only an academic pursuit but also of great importance in terms of being central to its effective conservation and management. Therefore, this research aimed to examine dietary composition, ontogenetic variations, and seasonal variations in the feeding behavior of L. intermedius in the Beles River. Area-specific information provides a foundation for developing targeted measures that could be aimed at ensuring that this valuable fisheries resource in the dynamic Blue Nile basin ecosystem is harvested sustainably.
Materials and Methods
The study was conducted in the Beles River of Mandura District in the Metekel Zone of the Benishangul-Gumuz Regional State, Ethiopia. The sampling area is located 50 km to the west of Gelgel Beles Town, which is the capital of Metekel Zone, at 11º 00’ºN latitude and 35º 45’ºE longitude. The Beles River flows around Gilgel Beles Town and Mankush (formerly Guba) District in Babizenda Kebele, 541 km and 696 km northwest of Addis Ababa, respectively (Fig. 1).
The shape of the catchments of the Beles River is elongated along the main course of the river, broadening towards the southwest as more tributary streams join the main river. The Beles River is fed by many rivers before it joins the River Abay (Blue Nile River), which is one of the largest rivers and also the river that makes the GERD. Abat Beles, Gilgel Beles, Durra, Ardi, Alaltu, Dondor, Qarsa, and Shar Rivers are the most important tributaries of the Beles River.
In the Beles River, there are 22 fish species belonging to 5 orders, 7 families, and 13 genera. Among the families, the Cyprinidae family comprised 10 species; L. intermedius is the dominant species among the family members (Getahun et al., 2020). According to Berie’s (2007) report, different types of phytoplankton that are found in the Beles River include Cyanophyceae (cyanobacteria), Chlorophyceae (green algae), Bacillariophyceae (Diatoms), Dinophyceae (Dinoflagellates), Cryptophyceae (Cryptophyta), and Euglenophyceae (Euglenophyta). The river is basically dominated by large populations of rotifers, copepods, cladocerans, ostracods, and dipteran insects. The Ciliophorans, especially Paramecium, were also represented by quite a number of species (Berie, 2007).
Fish samples were purchased from the landings of the fishers for eight consecutive months (January to September 2023) consecutive months, including four during the dry season and four during the rainy season. Total length (TL), fork length (FL), and standard length (SL) were measured to the nearest 0.1 centimeter. The total weight (TW) was measured to the nearest 1 g. Gut contents of sampled fish were removed and preserved in a 5% formalin solution (Dadebo et al., 2013). The 5% formalin inhibits the decomposition of the samples.
The gut contents were examined, and prey items were identified. Large food items, especially macrophytes (fruit), were identified visually; whereas, a dissecting microscope (MS5, magnification 10×; Leica, Wetzlar, Germany) and a compound microscope (magnification 100×; Leica DME, Wetzlar, Germany) were used to identify microscopic food items (Ademasu & Dadebo, 1997; Admassu et al., 2015; Dadebo et al., 2013; Desta et al., 2006). The relative importance and contribution of each food item to the diet of L. intermedius were determined using the standard methods, i.e., the frequency of occurrence method and volumetric analysis (Hyslop, 1980).
The number of gut samples containing one or more of a given food item was expressed as a percentage of all non-empty guts examined from dissected fish (Bagenal & Braum, 1978). The proportion of the population that feeds on certain food items was estimated by this method as follows.
Each prey was classified into different taxonomic categories, and the volume of each prey item was measured. The volumetric percentage of each prey organism was then estimated visually, considering their size and abundance relative to the total food items in the sample droplet. The volume of each food category was expressed as a percentage of the total food categories present in the gut content sample, following the following equation. The importance of different food categories in relation to the size of fish was determined by categorizing the TL of the fish into different size classes and by observing the volumetric contribution of different food items to each size class (Bowen, 1983).
Ontogenetic dietary shifts in L. intermedius were evaluated by quantifying the relative volumetric contribution of prey items across distinct size classes. Fish were grouped into eight total-length categories at 5 cm intervals, which were further consolidated into three broader classes: small (< 25 cm TL), medium (25–35 cm TL), and large (> 35 cm TL); to reduce misclassification, following the protocol of Dadebo et al. (2013). For each size category, the mean percentage volume of each prey type was calculated relative to the total gut content volume. These mean volumetric percentages served to illustrate how dietary composition changes as the fish grow.
Results
A total of 532 gut samples were analyzed, of which 75 (14.1%) specimens were found to be empty. The remaining 457 samples (85.9%) contained food items of both plant and animal origin. The plant-derived components included macrophytes, phytoplankton, and detritus, while animal-derived contents comprised insects, zooplankton, and gastropods. Among the insect prey, identifiable taxa included Chironomidae larvae, Trichoptera, Anisoptera, and Ephemeroptera.
The frequency of occurrence showed that detritus and phytoplankton were found to be the most important food items, occurring in 90.6% and 87.7% of the guts, respectively (Table 1). The high frequency of occurrence of phytoplankton was mainly due to green algae. The others, next to detritus and phytoplankton, were macrophytes (fruit, bark, and root), followed by insects and gastropods occurring in 52.4%, 37.7%, and 30.2% of the guts examined, respectively, and zooplanktons (5.8%) were less important than others in number. The high frequency of insects was due to Chironomidae larvae, followed by Ephemeroptera, and Anisoptera and Tricoptera, respectively, and the high frequency of gastropods was due to Mellanoides and Bullinus.
Volumetrically, detritus was the most important food item, constituting 42.1% of the total volume, followed by phytoplankton with 41.5% of the total volume of the gut contents, respectively. Macrophytes (7.2%), insects (4.3%), zooplankton (3.4%), and gastropods (1.5%) constituted relatively low volumes compared to the above food categories (Table 1). Food items that contain insects, Tricoptera, and Anisoptera in high volume. The Gastropods (1.5%) were mainly contained in Mellanoides and Bullinus.
There was no significant variation in the types of food consumed by L. intermedius between the dry and wet months, although a slight difference was observed in the volumetric proportions of the diet. Overall, except for insects and detritus, the contributions of all food categories were slightly higher during the wet season. In contrast, the volumetric contributions of insects and detritus were greater during the dry season. Additionally, except for detritus and zooplankton, the frequency of occurrence of all food items increased during the wet months (Table 2).
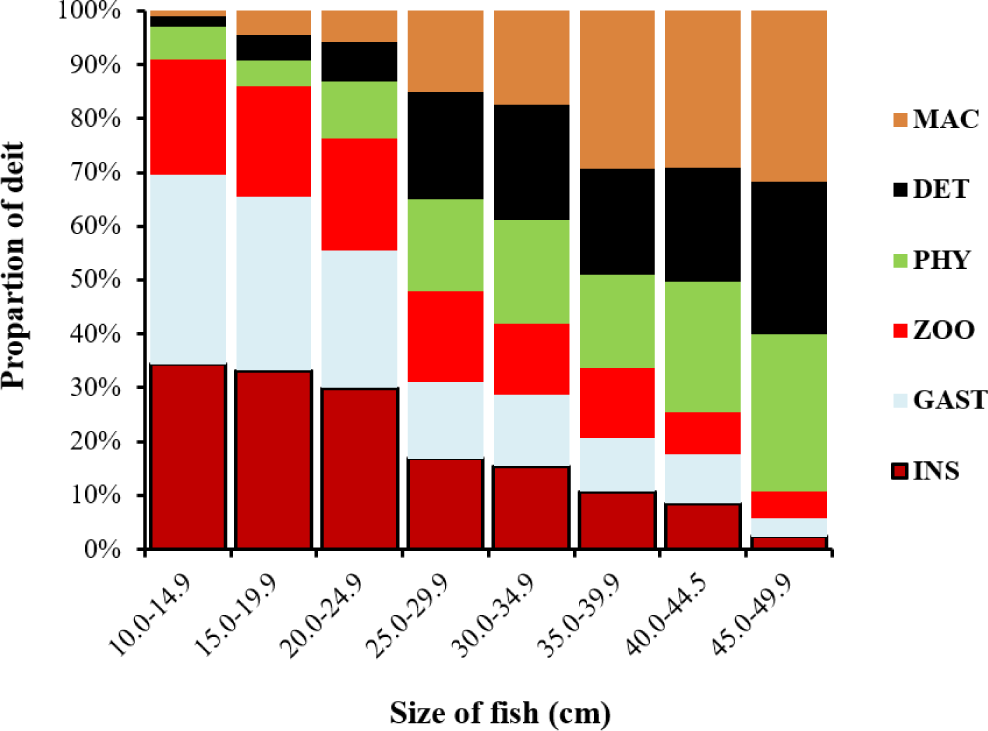
The TL of L. intermedius specimens containing food items ranged from 11.6 cm to 49.3 cm. To facilitate analysis, individuals were categorized into eight 5-cm length classes. These classes were further consolidated into three broad size categories: small (< 25 cm TL), medium (25–35 cm TL), and large (> 35 cm TL). Significant ontogenetic shifts in dietary composition were observed across these size classes. The volumetric contribution of insects, gastropods, and zooplankton decreased markedly with increasing fish size. Insects constituted over 50% of the diet in individuals smaller than 15 cm TL. Conversely, consumption of other food categories, phytoplankton, detritus, and macrophytes, increased proportionally with size. Gastropods were also predominantly consumed by smaller fish (Fig. 2). This indicates a distinct dietary transition towards phytoplankton and macrophytes during growth.
Quantitative diet analysis revealed the following volumetric contributions by size class:
According to Fig. 2, small (< 25 cm TL): insects (28.5%), gastropods (26.8%), zooplankton (19.9%), phytoplankton (9.6%), detritus (8.5%), macrophytes (6.6%). Medium (25–35 cm TL): detritus (20.7%), phytoplankton (18.2%), macrophytes (16.3%), insects (16.1%), zooplankton (15.1%), gastropods (13.7%). Large (> 35 cm TL): macrophytes (30.1%), phytoplankton (23.5%), detritus (23.1%), zooplankton (8.6%), gastropods (7.5%), insects (7.2%) (Fig. 2).
Discussion
Of the 532 fish gut samples analyzed, 75 specimens (14.1%) were empty. This phenomenon may be attributed to prolonged retention in fishing gear before collection by fishers and subsequent transport to markets. Extended capture times can facilitate gastric evacuation, regurgitation, or digestion of prey items, leading to an increased incidence of empty guts (Thormar et al., 2024). Based on the gut content analysis conducted in the present study, detritus, phytoplankton, macrophytes, insects, gastropods, and zooplankton were identified as significant food items, with their importance ranked from most to least dominant. Consistent with these findings, Dadebo et al. (2013) reported that L. intermedius in Lake Koka primarily feeds on detritus, phytoplankton, macrophytes, and insects. Similarly, another study has noted that the main food items of L. intermedius in Lake Koka include macrophytes, detritus, and aquatic insects (Kassahun, 2005).
Similarly, in Lake Tana, the diet of L. intermedius was dominated by benthic prey organisms, particularly insect larvae and detritus (de Graaf, 2003; Sibbing, 1998). Engdaw (2014) also reported a balanced intake of food items from both plant and animal sources. Kassahun (2005) and Dadebo et al. (2013) documented findings largely similar to those of the present study. However, the present study recorded fewer food categories than those reported by Kassahun (2005) and Dadebo et al. (2013). This difference may be attributable to variations in the sampling ecosystems, variations in the size classes of fish sampled, and the shorter sampling period in the present study. The divergent dietary compositions of L. intermedius across water bodies could be primarily driven by fundamental differences in habitat characteristics and prey communities. The evolution of piscivory is facilitated in Lake Tana due to its large size, deep depth with a well-established pelagic zone, and abundant populations of small endemic barb species (Nagelkerke, 1997). In contrast, the shallower and more turbid lake, Lake Koka, or other riverine environments often lack these specific prey fish but support high biomass of benthic invertebrates and detritus, favoring an omnivorous strategy (Dadebo et al., 2014). Furthermore, agricultural runoff can eutrophy water bodies, shifting primary production towards phytoplankton and detritus, thereby further shaping the available forage base and subsequently the diet of opportunistic feeders like L. intermedius.
However, contrary to the present investigation, the findings of Admassu & Dadebo (1997) and Zerihun (2020) indicated that the primary dietary sources of L. intermedius were of animal origin and detritus, with macrophytes, phytoplankton, zooplankton, and ostracods being less important. These studies also reported that the fish exhibited piscivorous tendencies above 30 cm TL. Furthermore, Desta et al. (2006), Nagelkerke (1997), and Wakijira (2013) documented piscivorous feeding habits in L. intermedius, with diets including fish, ostracods, zooplankton, mollusks, aquatic insects, and algae. This represents a different dietary composition compared to the present study. Specifically, Nagelkerke (1997) noted that in Lake Tana, Ethiopia, five of eight large barb species were specialized piscivores, attributing this behavior to the abundance of Barbus humilis in littoral areas and Barbus tanapelagius in the pelagic zone. Similarly, Desta et al. (2006) reported that L. intermedius in Lake Hawassa was piscivorous, primarily preying on the small barb Barbus paludinosus due to its abundance in the littoral zone and the absence of other specialized piscivorous fish. The reported inconsistencies in the published diet of L. intermedius are not unusual but indicative of the high trophic plasticity of the species. The inconsistencies can largely be attributed to spatial and temporal differences in the occurrence of prey items, ontogenetic variations, and variations in methodology in the diet analysis (Desta et al., 2006; Hyslop, 1980; Nagelkerke, 1997).
The significant volumetric contribution and frequent occurrence of detritus in the guts of L. intermedius sampled in this study highlight its importance as a food source. While detritus is recognized as having low nutritional value (Bagenal & Braum, 1978; Bowen, 1979), its increased consumption in lotic and lentic ecosystems often indicates scarcity of more nutritious primary foods (Bowen, 1979; King et al., 2003). Therefore, the reliance on detritus observed here implies a decline in the availability of higher-quality food resources within the Beles River.
As a consequence, this strong reliance on detritus detected in this research indicates reduced availability of higher-quality prey items (e.g., zooplankton, insects) in the Beles River. This shift in diet to one of reduced quality might be related to several ecological and anthropogenic factors. The Beles catchment is now under significant anthropogenic pressures, such as deforestation and agricultural encroachment (e.g., maize and sesame farming), that can lead to increased erosion and sedimentation (Berie, 2007). Such intensified sediment loads could exacerbate benthic habitats, suffocating gastropod and insect larval communities (King et al., 2003). In addition, runoff associated with farming is sure to carry fertilizers and pesticides into the riverine ecosystem. Fertilizers could augment phytoplankton biomass (perhaps explaining its high volumetric contribution), while pesticides might impact negatively upon zooplankton and macroinvertebrate numbers, because these are more sensitive toxins (Dadebo et al., 2013). Such possible factors are in accord with detected dietary habits and in line with documented impacts of land use changes upon global riverine food webs. This, therefore, indicates that L. intermedius trophic dynamics in the Beles River might act as a bioindicator of wider ecosystem disruption.
High ingestion of detritus, a resource usually having low nutritional value and high quantities of indigestible material (Bowen, 1979), has important ecological connotations for L. intermedius populations. Strong obligatoriness of detritivory usually requires a high ingestion rate in compensation for low assimilation of nutrients, hence high gut fullness but potentially low growth rate and reduced body condition (i.e., low weight-for-length) in comparison to populations whose diets are comprised of more protein-enriched sources like animal prey or water-column insects (Dadebo et al., 2013). The feeding strategy comes at a trade-off: while detritus is a ubiquitous and reliable resource base, resource consumption tends to define the maximum possible size and reproductive potential of individuals in the populations.
However, the feeding on detritus represents a major adaptive strategy promoting stable populations. In communities defined by the episodic or unpredictable nature of prey (such as seasonal insect emergences and fluctuating fish stocks), the exploitation of detritus represents a necessary ‘fallback’ resource. This allows L. intermedius to maintain a sustainable population density even without readily preferred, high-quality prey and hence augments the species’ resilience and reduces the risk of catastrophic population declines (Zerihun, 2020). Consequently, the detritivory represented by this research very probably reflects a strategy geared more towards the persistence and stability of populations rather than the optimization of individual growth performance, a common adaptation in resource-limited or fluctuating freshwater ecosystems.
This study aimed to investigate potential differences in the diet of L. intermedius between the dry and wet sampling months. Our findings revealed slight differences in variation in either the types of food items consumed or their volumetric contributions between the two seasons. In contrast, Admassu & Dadebo (1997) reported a clear seasonal pattern, with insects, macrophytes, fish, and gastropods contributing significantly to the diet during the dry season, while insects dominated during the wet season. Similarly, Dadebo et al. (2013) observed a pronounced seasonal difference in the diet of L. intermedius in Lake Koka. Their study showed that detritus (34.0%) and insects (32.5%) made up the majority of the diet in the dry season, whereas macrophytes (66.1%) and detritus (24.0%) were predominant in the wet season. In the Gilgel Gibe Reservoir, Wakijira (2013) also reported that aquatic insects, detritus, and zooplankton were more abundant in the diet during the dry season, whereas macrophytes dominated during the wet season.
An ontogenetic dietary shift was observed in the diet of L. intermedius in the Beles River. Individuals less than 25 cm in TL primarily consumed insects (28.5%), gastropods (26.8%), and zooplankton (19.9%). In contrast, fish in the 25–34.9 cm TL range exhibited a more balanced diet, with insects (16.1%), gastropods (13.7%), zooplankton (15.1%), phytoplankton (18.2%), macrophytes (16.25%), and detritus (20.65%) contributing relatively equally to their overall diet. For individuals exceeding 35 cm TL, detritus (23.1%), phytoplankton (23.5%), and macrophytes (30.1%) were the dominant dietary components. These findings suggest that dietary preference in L. intermedius is influenced by mouth size and the fish’s ability to process different food items efficiently. Consistent with the present study, Admassu & Dadebo (1997) documented distinct ontogenetic dietary shifts in L. intermedius from Lake Hawassa. They reported smaller fish (< 25 cm TL) primarily consumed insects and gastropods, while intermediate-sized fish (25–32 cm TL) fed approximately equally on insects, fish, gastropods, and macrophytes. Larger individuals (> 32 cm TL) were exclusively piscivorous, predominantly preying on Oreochromis niloticus. Similarly, Dadebo et al. (2013) observed pronounced ontogenetic dietary variation in Lake Koka: fish < 20 cm TL derived > 75% of their diet from detritus and insects, whereas those > 20 cm TL shifted to diets dominated (> 75% by volume) by macrophytes and detritus. This transition reflected declining insectivory and increasing herbivory with fish size (Dadebo et al., 2013). These dietary shifts reduce intraspecific competition between size classes. As juveniles and adults target different prey, the population can more efficiently partition available resources, which supports higher overall population density and stability within the ecosystem.
Contrary to our findings, studies of L. intermedius in Lake Hawassa detected negligible dietary differences across size classes. Desta et al. (2006) found no significant ontogenetic variation, though smaller fish (< 15 cm TL) consumed slightly more insects, fish eggs, and ostracods, while larger fish (> 25 cm TL) consumed more gastropods, fish, and detritus. Zerihun (2020) corroborated this pattern, noting higher insect consumption in fish < 25 cm TL and increased gastropod and macrophyte intake in larger size classes (> 25 cm TL).
Conclusion
The study reveals that L. intermedius in the Beles River is primarily detritivorous-phytoplanktivorous, with detritus (42.1%) and phytoplankton (41.5%) dominating its diet. Notable ontogenetic dietary shifts occur: smaller fish (< 25 cm TL) rely heavily on insects, gastropods, and zooplankton, while larger individuals (> 35 cm TL) transition to macrophytes, phytoplankton, and detritus. No significant seasonal variation was observed, indicating consistent resource utilization year-round. The dominance of low-nutritional-value detritus suggests reduced availability of high-quality prey, possibly reflecting ecological stress in the Beles River ecosystem. These findings highlight the species’ adaptive trophic plasticity but underscore potential resource limitations in this critical tributary of the Blue Nile basin.
Recommendations
Based on the results of the current investigation, the following points are recommended.
-
Conservation measures: protect riparian vegetation and benthic habitats to maintain detritus and phytoplankton production, ensuring food security for juvenile and adult L. intermedius.
-
Fisheries management: regulate fishing pressure on smaller size classes (< 25 cm TL), which exhibit higher vulnerability due to their specialized insect-based diet.
-
Long-term monitoring: extend sampling duration to validate seasonal stability and assess impacts of climate or anthropogenic changes on food resources.
-
Habitat restoration: investigate causes of reduced prey diversity (e.g., pollution, flow alterations) and restore degraded stretches to enhance prey availability.
-
Comparative studies: expand research to other Blue Nile tributaries to evaluate regional dietary plasticity and inform basin-wide conservation strategies.