






Introduction
Ethiopia has an extensive freshwater resource consisting of lakes, rivers, reservoirs and wetlands (Tesfaye & Wolff, 2014). The country is characterized by 12 major basins, eight of which are riverine, one lake and three dry basins (Tareke & Awoke, 2022).
The Awash and Omo-Gibe Rivers drain into Lakes Abbe and Turkana, which are shared by Ethiopia with Djibouti and Kenya, respectively. The Baro-Akobo Basin connects to the White Nile. The Abbay (Blue Nile) Basin rises from Lake Tana and forms the great Nile River that stretches through Egypt and empties into the Mediterranean Sea. These Ethiopia’s fluvial ecosystems are homes to rich and endemic ichthyofauna (Habteselassie, 2012; Lakew et al., 2016; Wakjira & Getahun, 2017).
Ethiopian rivers are important fish habitats (Wendimu et al., 2024). The extensive river networks harbour a diverse array of endemic and ecologically and socio-economically important fish species. Fish assemblage diversity and abundance in Ethiopia are influenced by various river systems including geography, hydrology and human activities (Lakew et al., 2016). The study of fish assemblages in Ethiopian rivers is an important ecological research, particularly in the face of increasing anthropogenic pressures, yet trustworthy comprehensive biological studies of these ecosystems are scarce in Ethiopia (Eyayu & Getahun, 2022a). Recent studies highlight the need to understand ecological and biological aspects of fish assemblages for effective management and conservation (Eyayu & Getahun, 2022a, 2024; Tecklie, 2024).
Although numerous studies have examined specific aspects of Ethiopian riverine fishes, a systematic synthesis of ecological patterns, biological traits, and management implications remains lacking. This review addresses this gap by compiling and analyzing published literature on riverine fish assemblages in Ethiopian river, with particular emphasis on diversity, distribution, habitat associations, biological characteristics, trophic ecology, major anthropogenic threats, conservation gaps, and management implications. By integrating available evidence, this review aims to provide a scientific foundation for sustainable riverine fisheries management and conservation planning in Ethiopia.
Materials and Methods
This review was conducted following a structured systematic review framework to ensure transparency, reproducibility, and methodological rigor. The approach systematically identified, screened, and synthesized published and grey literature addressing the ecological and biological characteristics of riverine fish assemblages in Ethiopia and their implications for sustainable fisheries management and conservation.
A comprehensive literature search was conducted across four major databases: Web of Science, Scopus, Google Scholar, and Google. Additional grey literature was retrieved from Ethiopian university repositories and institutional archives. Searches were performed using combinations of keywords and Boolean operators, including: “riverine fish assemblages,” “river fish diversity,” “freshwater fish ecology,” “fish biology,” “feeding ecology,” “reproductive biology,” “fisheries management,” and “Ethiopia.”
The search covered studies published between 2011 and 2025, emphasis on studies published from 2014 onwards. Reference lists of selected articles were further screened to identify additional relevant studies.
Studies were included if they met the following criteria: (i) focused on riverine or stream ecosystems within Ethiopia; (ii) examined ecological, biological, or assemblage-level attributes of freshwater fishes (e.g., species composition, diversity, abundance, growth, reproduction, or feeding ecology); and (iii) were published in English as peer-reviewed articles, academic theses, books, workshop proceedings, or reputable institutional reports.
Studies were excluded if they focused exclusively on lakes, reservoirs, marine systems, or aquaculture without explicit relevance to riverine environments, or if they lacked primary ecological or biological data relevant to fish assemblages.
All retrieved records were compiled and duplicates removed. Titles and abstracts were screened to exclude irrelevant studies, followed by full-text assessment based on the predefined inclusion criteria. After screening, a total of 100 studies were retained for qualitative synthesis. (Fig. 1). A total of 420 records were initially identified through database searches and other sources. After removal of 70 duplicate records, 350 unique records remained for title and abstract screening. Of these, 200 records were excluded as irrelevant, and 150 full-text articles were assessed for eligibility. Following full-text evaluation, 50 articles were excluded due to lack of riverine focus or insufficient ecological data. Ultimately, 100 studies were included in the final qualitative synthesis.
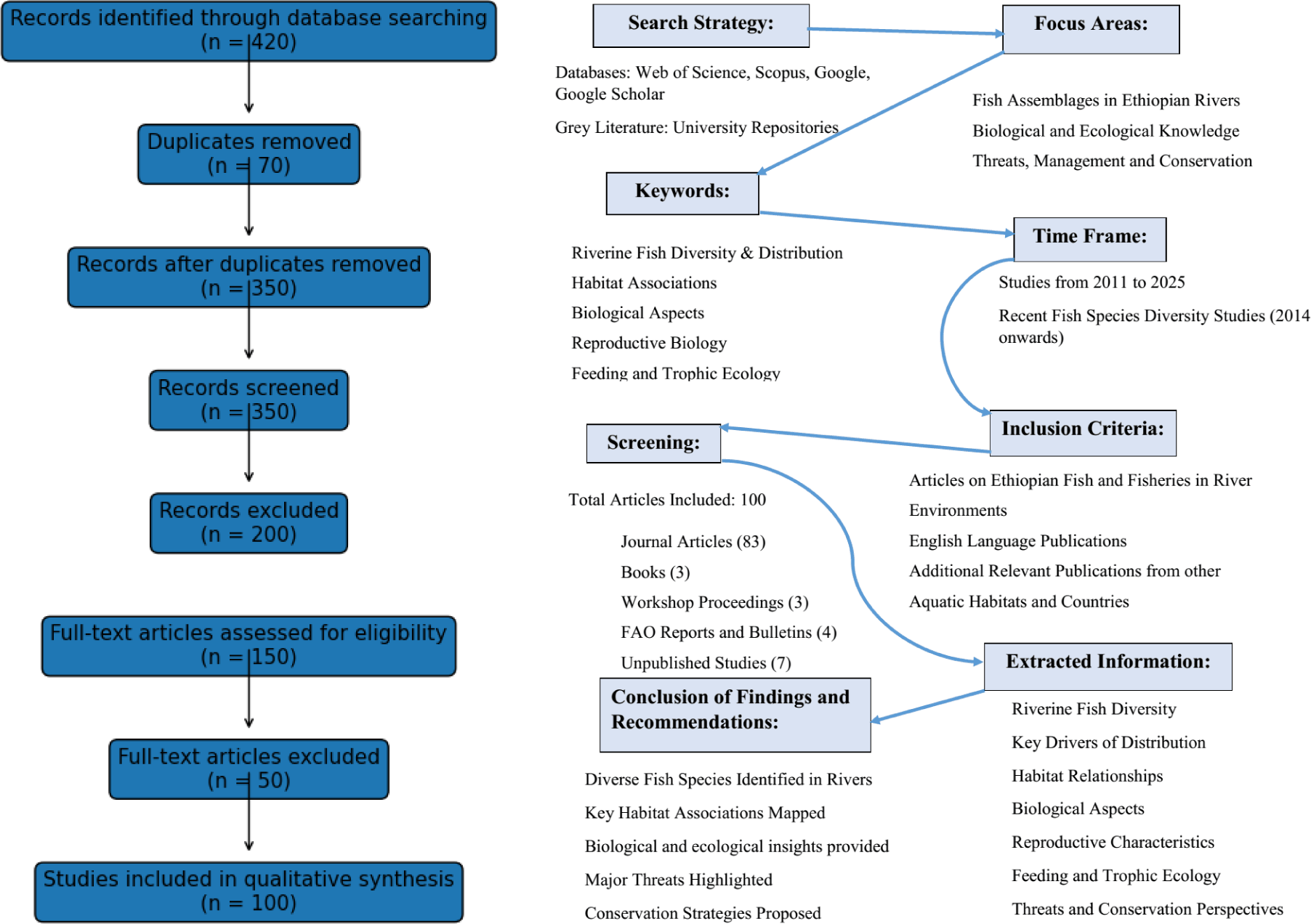
From each included study, information was extracted on study location, river basin, sampling design, fish species composition, assemblage structure, habitat characteristics, biological parameters, and reported anthropogenic pressures. Due to heterogeneity in study objectives, methodologies, and reported metrics, a qualitative synthesis approach was adopted. The extracted information was organized thematically to identify consistent ecological patterns, biological traits, threats, and management-relevant insights across Ethiopian river systems.
Results and Discussion
Ethiopia, the ‘water tower’ of East Africa, has a diverse ichthyofauna in its lakes, rivers and reservoirs (Getahun, 2017). The country has more than 200 fish species ascribed in to 75 genera, 31 families and 12 orders (Habteselassie, 2012). Of these, 40 are endemic and 11 are introduced (Getahun, 2017). Most introduced species have been introduced to water bodies in central and southeast Ethiopia. The Rift Valley lakes has the highest number of introduced species (Tesfaye & Wolff, 2014).
Ethiopian freshwater fishes fall into three categories: the Nilo-Sudanic, East African and endemic affinities. Nilo-Sudanic groups include the genera Alestes, Bagrus, Barilius, Citharinus, Hydrocynus, Hyperopisus, Labeo, Malapterurus and Mormyrus. These groups have similarities with species of West Africa and are considerably dominant in the Baro-Akobo, Webi-Shebele, Omo-Gibe and Nile Basin. The East African highland forms relate to eastern and southern Africa and parts of the Arabian Peninsula, and include the genera Clarias, Garra and Oreochromis. They occur in the northern and central Rift Valley Lakes, the Awash Basin, Highland Lakes (Tana and Hayq) and tributaries of the Abbay Basin. The endemic fish species are thought to exist nationwide (Wendimu et al., 2024).
Quantitative studies indicate that rivers and streams host more fish species than lakes and reservoirs (Temesgen et al., 2021; Wendimu et al., 2024). Among Ethiopian river basins, species richness is highest in Baro-Akobo (113), followed by Abbay (Blue Nile) (77) and Omo-Gibe (76–79) (Fig. 2) (Getahun, 2017; Wendimu et al., 2024). Endemicity is particularly high in the Abbay Basin, with Lake Tana sub-basin having 28 species (20 endemic). This diversity reflects the “species flock” of Labeobarbus in Lake Tana and highland tributaries (Wendimu et al., 2024).
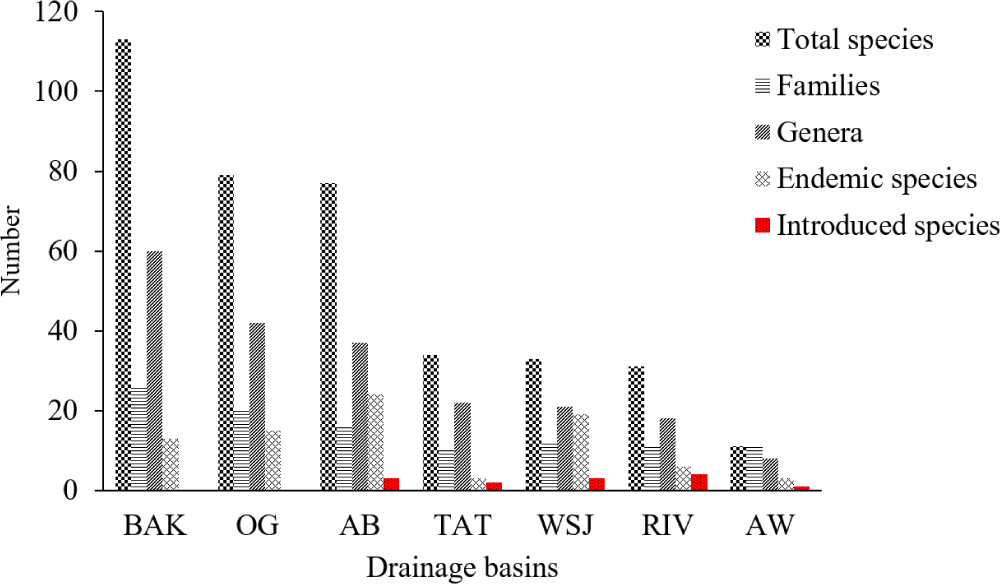
Fish assemblages demonstrate biogeographic structuring, with lowland systems supporting higher species richness due to habitat heterogeneity and connectivity. Uneven sampling and methodological differences underscore the need for standardized basin-wide surveys. However, the abundance and diversity of fish species are very uneven in the country’s drainage systems (Tesfaye & Wolff, 2014). Some recent studies investigated fish species diversity in different 28 rivers such as large, small and tributary rivers in their basin (Table 1). The studies revealed that Ethiopian riverine fish communities show significant differences, and the ecological importance of Ethiopian rivers as refugia of distinctive fish biodiversity (Gebremedhin et al., 2017).
| Name of river(s) | Location and basin | No. of species | Sources |
|---|---|---|---|
| Baro | Baro-Akobo Basin | 57 | Tesfaye & Wolff (2014) |
| Gelegu | Abbay Basin | 38 | Eyayu & Getahun (2024) |
| Ayima | Abbay Basin | 35 | Eyayu & Getahun (2024) |
| Gilo | Omo-Gibe Basin | 27 | Tut et al. (2019) |
| Omo | Omo-Gibe Basin | 26 | Wakjira & Getahun (2017) |
| Shinfa | Tekeze-Atbara Basin | 25 | Eyayu & Getahun (2024) |
| Beles and Gilgel Beles | Abbay Basin | 23 | Getahun et al. (2020) |
| Baro | Baro-Akobo Basin | 14 | Gebreselassie et al. (2021) |
| Kir | Baro-Akobo | 14 | Gebreselassie et al. (2021) |
| Gilgel Abay and Andassa | Abbay Basin | 13 | Aynalem et al. (2018) |
| Awash and its tributaries | Awash River Basin | 11 | Englmaier et al. (2020) |
| Gilgel Abbay | Abbay Basin | 10 | Aynalem & Mingist (2020) |
| Gumara | Abbay Basin | 10 | Erarto et al. (2020) |
| Gibe and Wabe | Omo-Gibe Basin | 10 | Mekonen & Hailu (2021) |
| Gerado | Abbay Basin | 10 | Tessema & Mohamed (2016) |
| Geba | Baro-Akobo | 9 | Melaku et al. (2017) |
| Sor | Baro-Akobo | 9 | Melaku et al. (2017) |
| Abbay below Tiss Issat Fall | Abbay Basin | 8 | Awoke et al. (2015a) |
| Andassa | Abbay Basin | 7 | Aynalem & Mingist (2020) |
| Gojeb | Omo-Gibe Basin | 7 | Mohammed (2014) |
| Drima | Awash River Basin | 6 | Tessema & Mohamed (2016) |
| Terie River | Abbay Basin | 6 | Tecklie (2024) |
| Jemma | Abbay Basin | 5 | Aynalem & Mingist (2020) |
| Koga | Abbay Basin | 5 | Aynalem & Mingist (2020) |
| Aveya | Abbay Basin | 4 | Mingist & Gebremedhin (2014) |
| Gilgel Gibe | Omo-Gibe Basin | 3 | Mohammed (2014) |
| Upper Awash River | Awash River Basin | 3 | Temesgen et al. (2021) |
Ethiopian fish diversity is shaped by geography, topography, geology, and a wide elevation gradient (Billi, 2015; Fashing et al., 2022; Van den Hende et al., 2021). Fish occur across desert-like lakes (e.g., Lake Afambo) and highland rivers (e.g., Woyib in the Bale highland), creating habitat heterogeneity. Environmental gradients, stream characteristics, human activity, and species interactions drive uneven fish distribution (Vijverberg et al., 2012; Wakjira & Getahun, 2017). For example, the Awash River shows distinct highland and rift valley communities separated by transition zones with high species diversity (Englmaier et al., 2020).
Altitude differences are mainly responsible for structuring fish communities in major river basins (Vijverberg et al., 2012; Wakjira & Getahun, 2017). The geographical diversity, ranging from highland plateaus to deep rift valleys, affects hydrology, substrate and nutrients in streams and hence associated ichthyofaunal diversities. More particularly, fish populations in Ethiopian river systems are influenced by habitat structure, water quality and seasonal changes (Abebe et al., 2021; Eyayu & Getahun, 2022a). Families such as cyprinidae, cichlidae and clariidae dominate Ethiopian fish assemblages owing to their ecological adaptations towards floodplain habitats (Englmaier et al., 2020).
Lacustrine fish fauna in Ethiopia are largely dependent on riverine tributaries (Gebru et al., 2019; Mequanent et al., 2022; Wakjira & Getahun, 2017). In Ethiopia, standing water dynamics are similar to rivers (Gebreselassie et al., 2021; Tesfaye & Wolff, 2014), and understanding lake–river dynamics and connectivity is critical for conservation and management (Wakjira & Getahun, 2017). For example, Labeobarbus species from Lake Tana migrate to rivers such as the Ribb for spawning, which is critical for survival, reproduction and genetic diversity (Anteneh et al., 2013; Gebremedhin et al., 2017). Assemblage variations often result from climatic zones, elevation gradients and river basin connectivity. Understanding these variations is therefore critical for developing effective management strategies to protect riverine biodiversity affected by brutal human activities.
Biological, chemical, and physical factors shape fish communities across spatial scales (Eyayu & Getahun, 2022a). Factors structuring fish communities in natural ecosystems are important for management and conservation such ecosystems (Eyayu & Getahun, 2022a; Tesfaye et al., 2016). Environmental variables such as temperature, flow, substrate, sediment, dissolved oxygen (DO), habitat complexity, and nutrients influence species composition and abundance (Wakjira & Getahun, 2017; Wendimu et al., 2024). The hydrology of the river plays a major role in the composition of fish community. Fish species composition and abundance differed in Ayima, Gelegu and Shinfa Rivers (Eyayu & Getahun, 2022a). Tecklie (2024) reported higher fish species diversity in deeper, rocky sites of the Terie River than in shallow, muddy substratum. Higher fish species in deeper and rockier sites lend fishes more suitable refuge sites (hiding under rocks) and deeper gorges assure fishes escape from incidence of predation and fishing than shallow and muddy habitats.
Water quality parameters like those that DO, pH and turbidity are strongly correlated with endemic Labeobarbus diversity in the Gumara River (Lake Tana’s tributary) (Abebe et al., 2021). This is because Labeobarbus species require well oxygenated water and gravel beds for spawning and reproduction (Teshome et al., 2015a, 2015b). DO, length of river channel, site depth, water transparency and nutrients largely influence floodplain river fish communities in the Alitash National Park (Eyayu & Getahun, 2022b). In the Gumara River, temperature, water velocity and substrate types affect abundance of fish species (Erarto et al., 2020; Zelalem et al., 2022).
Different fish communities are supported in different river habitats, including riffles, pools and backwaters. Tesfay et al. (2019) observed that, riffles are the least occupied and pools and runs have the highest fish diversity and species richness. Wubie et al. (2017) and Temesgen et al. (2021) reported that most fish were from pool units in the Upper Awash River because of its depth, low velocity and high water volume and serve as a refuge for ichthyofauna. Riffles have less water volume and fewer food resources, which limit species richness. Water temperature change, high flow velocity, turbidity, allochthonous nutrients and food availability restrict species diversity in riffles (Wubie et al., 2017). In Gumara River, pools had 2,160 individuals (9 species) vs. 1,266 in riffles and 454 in runs with low diversity and evenness, indicating species dominance and uneven distribution along a river courses (Erarto et al., 2020).
Fish species require a specific habitat in the river. Garra spp. prefer the gravel and boulder bed because gravel substrates protect eggs and juveniles from riffles wash aways (Erarto, 2019). Shitaw et al. (2018) noted Labeobarbus spp. favor fast-flowing, clear, well oxygenated water and gravel-bed streams. These preferences are critical for survival and reproduction in the natural environment. Changes in catchment area, canopy closure, substrate type, distance from headwater, river depth and width in Beles and Gilgel Beles Rivers were associated with upstream to downstream increases in species (Getahun et al., 2020). In Erarto (2019) study, abundance of Garra spp. decreased from upstream to river mouth due to sand, gravel and large rocks.
River habitat structure is important for fish species distribution because different units provide options for different species (Tesfay et al., 2019). The Beles River harbors more fish species than Gilgel Beles because of dense vegetation, a larger catchment area, and different substrate (Getahun et al., 2020). Habitat complexity increases fish diversity because areas such as submerged vegetation and rock structures provide refuges for many species (Eyayu & Getahun, 2022a). Anteneh et al. (2013) and Zelalem et al. (2022) pointed out that rivers are important for various life stages of fish, including spawning, nursery, and as feeding grounds. Fish abundance, distribution, and diversity reflect waterbody health (Gebru et al., 2019). In the Upper Awash River, water volume, pollution, and habitat modification cause drastic changes in fish species composition and abundance (Wubie et al., 2017).
Fish biology, including reproductive and feeding aspects, is fundamental to sustainable fisheries management (Tesfaye et al., 2016) and modeling aquatic ecosystems for developing management strategies, evaluating the impact of human activities on fish populations, and identifying potential threats (Temesgen et al., 2023). Research on fish reproductive biology is fundamental to conservation and management strategies (Muchlisin, 2014), and is key for selecting promising fish candidates for aquaculture (Muchlisin, 2013). Knowledge on fish feeding ecology is fundamental to managing and conserving populations and their habitats (Eyayu & Getahun, 2022b; Mingist et al., 2023). Research on fish diets is fundamental to fisheries and aquaculture management programs (Eyayu & Getahun, 2022b; Mingist et al., 2023; Wakjira, 2016). However, little information is available on the biology of fishes in Ethiopian rivers (Eyayu & Getahun, 2022b; Temesgen et al., 2023). Herein, we discuss the biology of fishes in Ethiopian rivers with a focus on their biological aspects, reproductive and feeding behaviors and highlight the importance of management and conservation.
Length–weight relationships (LWRs), condition factors (CFs), fecundity, and sex ratios (SRs) reflect environmental control on fish life-history traits in Ethiopian rivers. Variability within species across rivers suggests that hydrology, habitat quality, and anthropogenic disturbance often override intrinsic species-level traits. Consequently, river-specific biological reference points are more appropriate than generalized national values for fisheries assessment and management. Values summarized in Tables 2–5 should not be interpreted as directly comparable across rivers or studies because of substantial differences in sampling season, fishing gear selectivity, size ranges, and analytical methods. These tables are intended to illustrate the range of biological variation reported for Ethiopian riverine fishes and to identify research coverage and gaps, rather than to support quantitative cross-system comparisons.
LWRs are fundamental tools in fisheries biology and management (Bwathondi & Abdulkarim, 2017). LWRs enable the conversion of length-frequency data into biomass estimates, support stock assessment and yield estimation, and provide insights into fish condition, growth performance, and habitat quality (Asadi et al., 2017). LWR reflects fish condition and growth patterns where as weight increases, it is positive allometric growth (b > 3) for fatter fish and negative allometric growth (b between 2 and 3) for leaner fish. Isometric growth means weight increases proportionally to body length at constant specific gravity, while allometric growth does not (Asadi et al., 2017).
In Ethiopian rivers, LWR b-values vary spatially and interspecifically, indicating strong environmental influence (Table 2). Both isometric (b ≈ 3) and allometric growth patterns (b ≠ 3) are observed, and the same species frequently exhibits different b values across river systems. Such variability suggests that growth patterns are strongly influenced by environmental conditions rather than intrinsic species traits alone. Oreochromis niloticus had negative allometric growth with b values of 2–3 in both seasons and Labeobarbus intermedius had isometric (b > 3) and negative allometric (b < 3) growth in the Terie River (Tecklie, 2024). In the Gilo River, O. niloticus had a b-value that was lower than 3, indicating negative allometric growth (Tut et al., 2019). L. intermedius and Labeo forskalii had negative allometric growth in the Gilgel Gibe and Gojeb Rivers, indicating that there were differences in food availability or environmental conditions (Mohammed, 2014). From Table 2, it is clear that the same species of fish has different body growth types and b-values in different rivers.
| Fish species | LWR b-value | River(s) name | Sources |
|---|---|---|---|
| Bagrus bajad | 2.68 | Gojeb | Mohammed (2014) |
| Clarias gariepinus | 2.4 | Andassa | Aynalem & Mingist (2020) |
| 2.6 | Gilgel Abay | Aynalem & Mingist (2020) | |
| Garra dembeensis | 2.34 | Upper Awash River | Temesgen et al. (2023) |
| Garra quadrimaculata | 2.74 | Upper Awash River | Temesgen et al. (2023) |
| Labeo forskalii | 3.09 | Beles | Berie (2020) |
| 2.86 | Gilgel Beles | Berie (2020) | |
| 2.79 | Abbay River, below Tisisat Fall | Awoke et al. (2015b) | |
| 2.70 | Gojeb | Mohammed (2014) | |
| Labeo cylindricus | 2.56 | Geba | Melaku et al. (2017) |
| 2.88 | Sor | Melaku et al. (2017) | |
| Labeobarbus intermedius | 2.70 | Geba | Melaku et al. (2017) |
| 2.27 | Sor | Melaku et al. (2017) | |
| 3.0 | Andassa | Aynalem & Mingist (2020) | |
| 3.1 | Gilgel Abay | Aynalem & Mingist (2020) | |
| 3.22 | Abbay River, below Tisisat Fall | Awoke et al. (2015b) | |
| 3.24 | Beles | Berie (2020) | |
| 3.36 | Gilgel Beles | Berie (2020) | |
| 2.60 | Gilgel Gibe | Mohammed (2014) | |
| 2.77 | Gojeb | Mohammed (2014) | |
| Labeobarbus nedgia | 2.60 | Geba | Melaku et al. (2017) |
| 2.70 | Sor | Melaku et al. (2017) | |
| 3.1 | Andassa | Aynalem & Mingist (2020) | |
| 3.3 | Gilgel Abay | Aynalem & Mingist (2020) | |
| Mormyrus kannume | 2.59 | Abbay River, below Tisisat Fall | Awoke et al. (2015b) |
| Labeobarbus beso | 3.1 | Andassa | Aynalem & Mingist (2020) |
| 3.1 | Gilgel Abay | Aynalem & Mingist (2020) |
Differences in b-values among rivers likely reflect variation in food availability, hydrological regime, habitat structure, and anthropogenic disturbance. Rivers with stable flow, diverse substrates, and higher productivity tend to support isometric or positive allometric growth, whereas systems affected by high flow variability, sedimentation, water abstraction, or pollution often exhibit negative allometric growth. These patterns highlight the ecological relevance of LWRs as indicators of habitat quality and ecosystem condition.
However, interpretation of LWR parameters requires caution. Growth coefficients are influenced by several biological and methodological factors, including seasonality, maturity stage, sex, sample size, length range, and fishing gear selectivity. Seasonal changes in food availability and reproductive investment can substantially alter weight-length relationships, particularly in tropical rivers with pronounced wet–dry cycles. In addition, comparisons among studies should ideally consider confidence intervals for parameters a and b and coefficients of determination (R²), although these metrics are not consistently reported in all reviewed studies. Overall, the observed variability in LWR parameters among Ethiopian rivers underscores the need for river-specific growth models and locally informed fisheries management strategies. Applying generalized LWR parameters across basins may lead to biased biomass estimates and misinformed management decisions, particularly in heterogeneous riverine environments.
CF is a biometric criterion for the assessment of fish condition based on weight and length. Higher weights indicate healthier physiological states at a given length (Bwathondi & Abdulkarim, 2017). It is dependent on maturity, spawning, life cycle, environmental factors and food availability (Asadi et al., 2017). CF studies are important for fisheries management (Asadi et al., 2017; Muchlisin, 2014).
Table 3 showed the CF value variation among species and within species in Ethiopian rivers, influenced by seasonal, local, age, sampling time, food availability and gonad development (Tut et al., 2019). Higher values indicate better body condition with faster growth and environmental adaptation (Asadi et al., 2017). Environmental factors including food quality and quantity, water level, disease, affect growth and reproduction of fish. Higher condition indicates higher energy, higher food base, good reproductive potential or favorable conditions. Fishes in rivers have low CFs due to food quantity, quality, water level, flow rate and temperature (Awoke et al., 2015b; Berie, 2020). Males and females differ in fish CFs due to biological factors that affect growth and development. Teshome et al. (2015b) show females in L. intermedius and L. brevicephalus tributaries of Lake Tana have better CFs likely from smaller fork length and larger total weight from gonadal development.
| Fish species | Mean of CF | River(s) name | Sources |
|---|---|---|---|
| Bagrus bajad | 1.24 | Gojeb | Mohammed (2014) |
| Clarias gariepinus | 0.51 | Andassa | Aynalem & Mingist (2020) |
| 0.95 | Gilgel Abay | Aynalem & Mingist (2020) | |
| Garra dembeensis | 1.34 | Upper Awash River | Temesgen et al. (2023) |
| Garra quadrimaculata | 1.23 | Upper Awash River | Temesgen et al. (2023) |
| Labeo forskalii | 0.94 | Beles | Berie (2020) |
| 1.00 | Gilgel Beles | Berie (2020) | |
| 0.84 | Abbay River, below Tisisat Fall | Awoke et al. (2015b) | |
| 1.22 | Gojeb | Mohammed (2014) | |
| Labeobarbus brevicephalus | 1.21 | Tributary rivers of Lake Tana | Teshome et al. (2015b) |
| Labeo cylindricus | 1.21 | Geba | Melaku et al. (2017) |
| Labeobarbus intermedius | 1.21 | Geba | Melaku et al. (2017) |
| 0.81 | Andassa | Aynalem & Mingist (2020) | |
| 0.89 | Gilgel Abay | Aynalem & Mingist (2020) | |
| 0.99 | Abbay River, below Tisisat Fall | Awoke et al. (2015b) | |
| 1.10 | Beles | Berie (2020) | |
| 1.00 | Gilgel Beles | Berie (2020) | |
| 1.23 | Borkena | Tessema et al. (2012) | |
| 1.31 | Mille | Tessema et al. (2012) | |
| 1.20 | Gilgel Gibe | Mohammed (2014) | |
| 1.20 | Gojeb | Mohammed (2014) | |
| 1.30 | Tributary rivers of Lake Tana | Teshome et al. (2015b) | |
| Labeobarbus nedgia | 1.05 | Geba | Melaku et al. (2017) |
| 0.84 | Andassa | Aynalem & Mingist (2020) | |
| 0.86 | Gilgel Abay | Aynalem & Mingist (2020) | |
| Mormyrus kannume | 0.49 | Abbay River, below Tisisat Fall | Awoke et al. (2015b) |
| Labeobarbus beso | 1.33 | Andassa | Aynalem & Mingist (2020) |
| 1.00 | Gilgel Abay | Aynalem & Mingist (2020) |
The most common reproductive parameters of fish species include fertility, spawning frequency, egg and larval size and quality, and timing and duration of spawning (Dominguez-Petit et al., 2022). Studies have investigated fish reproductive biology in Ethiopian lakes and reservoirs (Berie, 2020; Gebremedhin et al., 2012; Tesfaye et al., 2016; Tesfahun, 2018), but little is known about river ecosystems. Here, we examined and summarized the fish reproductive biology elements such as SR, length at first maturity, fecundity and spawning season.
SR is an important indicator to assess reproductive viability of fish because it changes with life cycle events affecting males and females. SR defines population structure, assesses reproductive potential and estimates fish stock size, ensuring proportional exploitation of both sexes (Neves et al., 2020). In stable conditions without frequent vibrations SR is not always 1:1 (Dominguez-Petit et al., 2017). Monitoring SR allows fisheries managers to avoid overexploitation of one sex, leading to population declines and genetic bottlenecks. Studying SR allows to evaluate the health of fish populations and their resilience to environmental changes.
Fish SR was reported to be unbalanced in Ethiopian rivers, and it deviated from 1:1 (Table 4). It could be influenced by biological mechanisms such as maturity, mortality, and habitat preferences. Fish SR is likely affected by fish vulnerability to gears and this causes deviation from 1:1 (Gebremedhin et al., 2012). Berie (2020) mentioned that females are more vulnerable because of ovarian development. Differences in SR in tributaries of Lake Tana involve different maturity, mortality, migration time and rate (Teshome et al., 2015a). Anteneh et al. (2013) suggested that L. intermedius imbalance is associated with spawning habits and females stay longer in breeding areas that could explain the imbalance.
| Fish species | SR (F:M) | River(s) name | Sources |
|---|---|---|---|
| Bagrus bajad | 1:1.14 | Gojeb | Mohammed (2014) |
| Bagrus docmak | 1:0.33 | Abbay River, below Tisisat Fall | Awoke et al. (2015b) |
| Clarias gariepinus | 1:0.31 | Abbay River, below Tisisat Fall | Awoke et al. (2015b) |
| 1:0.65 | Andassa | Aynalem & Mingist (2020) | |
| 1:0.82 | Gilgel Abay | Aynalem & Mingist (2020) | |
| Garra dembeensis | 1.03:1 | Upper Awash River | Temesgen et al. (2023) |
| Garra quadrimaculata | 1.05:1 | Upper Awash River | Temesgen et al. (2023) |
| Labeo forskalii | 1:0.96 | Beles | Berie (2020) |
| 1:0.21 | Abbay River, below Tisisat Fall | Awoke et al. (2015b) | |
| 1:1.09 | Gojeb | Mohammed (2014) | |
| Labeobarbus brevicephalus | 1:0.18* | Andassa | Aynalem & Mingist (2020) |
| 1:0.27* | Gilgel Abay | Aynalem & Mingist (2020) | |
| 1:1.72* | Tributary rivers of Lake Tana | Teshome et al. (2015b) | |
| Labeobarbus crassibarbis | 1:0.15 | Abbay River, below Tisisat Fall | Awoke et al. (2015b) |
| 1:0.1* | Andassa | Aynalem & Mingist (2020) | |
| Labeobarbus intermedius | 1:0.37* | Andassa | Aynalem & Mingist (2020) |
| 1:0.34* | Gilgel Abay | Aynalem & Mingist (2020) | |
| 1:0.31 | Abbay River, below Tisisat Fall | Awoke et al. (2015b) | |
| 1:0.67 | Gilgel Beles | Berie (2020) | |
| 1:1.25 | Gilgel Gibe | Mohammed (2014) | |
| 1:0.59 | Gojeb | Mohammed (2014) | |
| 1:1.82* | Tributary rivers of Lake Tana | Teshome et al. (2015b) | |
| Labeobarbus nedgia | 1:0.2* | Andassa | Aynalem & Mingist (2020) |
| 1:0.43* | Gilgel Abay | Aynalem & Mingist (2020) | |
| 1:0.72 | Abbay River, below Tisisat Fall | Awoke et al. (2015b) | |
| 1:55* | Tributary rivers of Lake Tana | Teshome et al. (2015b) | |
| Labeobarbus megastoma | 1:1.5 | Tributary rivers of Lake Tana | Teshome et al. (2015b) |
| Labeobarbus truttiformis | 1:3 | Tributary rivers of Lake Tana | Teshome et al. (2015b) |
| Labeobarbus tsanensis | 1:1.5 | Tributary rivers of Lake Tana | Teshome et al. (2015b) |
| Mormyrus kannume | 1:0.96 | Abbay River, below Tisisat Fall | Awoke et al. (2015b) |
| Oreochromis niloticus | 1:0.50 | Abbay River, below Tisisat Fall | Awoke et al. (2015b) |
| Labeobarbus beso | 1:0.27* | Andassa | Aynalem & Mingist (2020) |
| 1:0.37* | Gilgel Abay | Aynalem & Mingist (2020) |
Fish reach reproductive maturity at different ages and lengths (Neves et al., 2020). Young fish recruitment is influenced by the number of eggs available from sexually mature fish (Muluye et al., 2016; Tesfahun, 2018). Using the wrong fishing gear depletes sexually mature fish and affects recruitment and ecosystems (Muluye et al., 2016). FAO et al. (2017) advises commercial catch after the minimum harvestable size, length at first maturity (L50) which defines fish life history, reproductive potential and mature fish size (Tesfaye et al., 2016). L50 varies among species and depends on environmental conditions, resources, fishing pressure and biotic interactions (Soares et al., 2020). L50 is important for fish stock management preventing the exploitation of juvenile fish and damage to spawning stock (Neves et al., 2020). It guides gear choice for capture fisheries and mesh sizes targeting mature fish (Karna et al., 2012). L50 values are important for sustainable fish stock management ensuring long-term viability and ecosystem balance. Understanding L50 helps develop conservation strategies for vulnerable species (Soares et al., 2020).
For example, the L50 of L. quadrimaculata is 9.4 cm for males and 9.2 cm for females in the Upper Awash River, which implies that females mature at a smaller size (Temesgen et al., 2023). L50 variation among fish populations is caused by water physical and chemical properties (Tesfahun, 2018) and overfishing pressure (Mohammed & Uraguchi, 2013). Fish L50 size depends on genes and their environment, as documented by Tesfaye et al. (2016).
Fish biology studies focus on spawning season and fecundity to predict recruitment variability and protect recruits (Dominguez-Petit et al., 2017; Temesgen et al., 2023; Tesfahun, 2019). Gonadosomatic index (GSI) is a useful tool for fisheries management to reveal reproductive status and potential spawning periods. GSI values can be used to monitor reproductive status and spawning periods over time of several fish species (Tesfahun, 2019). Higher GSI indicates earlier maturation, greater reproductive investment and viability (Neves et al., 2020). Development of gonads is important for successful reproduction and species health and survival.
Fecundity is the egg-laying capacity of fish and is an important trait to assess reproductive potential, life history and fishery management (Muchlisin, 2014). Differences in fecundity of the same fish in Ethiopian rivers (Table 5) could be attributed to genetic or habitat differences. Fecundity is associated with body length, weight and gonad weight (Admassu et al., 2015). Species like L. intermedius, L. nedgia and L. beso show strong positive correlations with total length, weight and gonad weight in Andassa and Gilgel Abay Rivers (Aynalem & Mingist, 2020). Berie (2020) found similar results: L. intermedius fecundity in Beles and Gilgel Beles Rivers was strongly correlated with gonad weight, body weight and length.
| Fish species | Mean fecundity | River(s) | Sources |
|---|---|---|---|
| Garra quadrimaculata | 691.69 | Upper Awash River | Temesgen et al. (2023) |
| Labeobarbus intermedius | 3,595 | Andassa | Aynalem & Mingist (2020) |
| 3,595 | Gilgel Abay | Aynalem & Mingist (2020) | |
| 3,705 | Abbay River, below Tisisat Fall | Awoke et al. (2015b) | |
| 4,208 | Beles | Berie (2020) | |
| Labeobarbus nedgia | 510 | Andassa | Aynalem & Mingist (2020) |
| 510 | Gilgel Abay | Aynalem & Mingist (2020) | |
| Labeobarbus beso | 5,529 | Andassa | Aynalem & Mingist (2020) |
| 5,529 | Gilgel Abay | Aynalem & Mingist (2020) |
Fish breeding seasons are estimated based on visual observations or by using the five-stage gonad maturity: immature (I), recovering spent or developing virgin (II), ripening (III), ripe (IV) and spent (V). The percentages of ripe gonads in each month define the breeding seasons, i.e., the period when most fish are in breeding condition (Dadebo et al., 2011). Tropical inland water bodies breed throughout the year but are influenced by rainfall related factors such as water level changes and seasonal flooding (Admassu et al., 2015). In Ethiopian water bodies, breeding is related to rainfall and periodicity with most spawning occurring during the rainy season when fish food is abundant (Muluye et al., 2016; Tesfahun, 2018). Ethiopian river fish species breed seasonally and are influenced by temperature and water flow cues shaped by seasonal rainfall patterns. For example, Labeobarbus species such as L. brevicephalus, L. intermedius, L. nedgia and Labeobarbus tsanensis migrated to Lake Tana’s Arno-Garno River in August to October (Gebremedhin et al., 2012). Labeobarbus species such as L. intermedius, L. brevicephalus, Labeobarbus platydorsus, L. truttiformis, L. tsanensis, L. surkis, L. gorgorensis, L. crassibarbis and L. nedgia have peak breeding from August–October in Lake Tana’s tributaries, including Zabzi, Kilty, Gilgel Abay and Jemma Rivers (Mequanent et al., 2014).
Effective management of breeding seasons is critical for Ethiopian fish resources. Regulations protect populations at vulnerable times, contributing to conservation and aquatic ecosystem health over the long term (Tesfahun, 2018). Monitoring peak breeding seasons ensures sustainable fish populations (Admassu et al., 2015).
Food and feeding are critical to fish physiology, ecology and health, impacting survival and health (Bakhtiyar et al., 2023). Food and trophic preferences influence growth and development because of complex life histories often changing during life (Bakhtiyar et al., 2023). Studies on fish feeding ecology in Ethiopian water bodies is increasing (Admassu et al., 2015; Dadebo et al., 2014a, 2014b; Temesgen et al., 2022; Tesfaye et al., 2020; Teshome et al., 2023; Wakjira, 2016), but that of riverine fishes is less known (Eyayu & Getahun, 2022b). Fishes of Ethiopian rivers have a wide range of feeding strategies (Teshome et al., 2023), from herbivores to top predators, highlighting the importance of fish assemblages for trophic dynamics and nutrient cycling. Different species in different environments may have different gut food items because of varying feeding habits. G. quadrimaculata and L. beso feed on phytoplankton, detritus, sand and zooplankton (Temesgen et al., 2023), whereas that of L. intermedius varied by season, prey, habitat and fish size (Mingist et al., 2023). Large cyprinid fishes in the upper Abbay River eat different food items frequently and volumetrically with seasons (Teshome et al., 2023). Eyayu & Getahun (2022b) noted that only some predatory fish preys varied in dietary importance due to season and size differences.
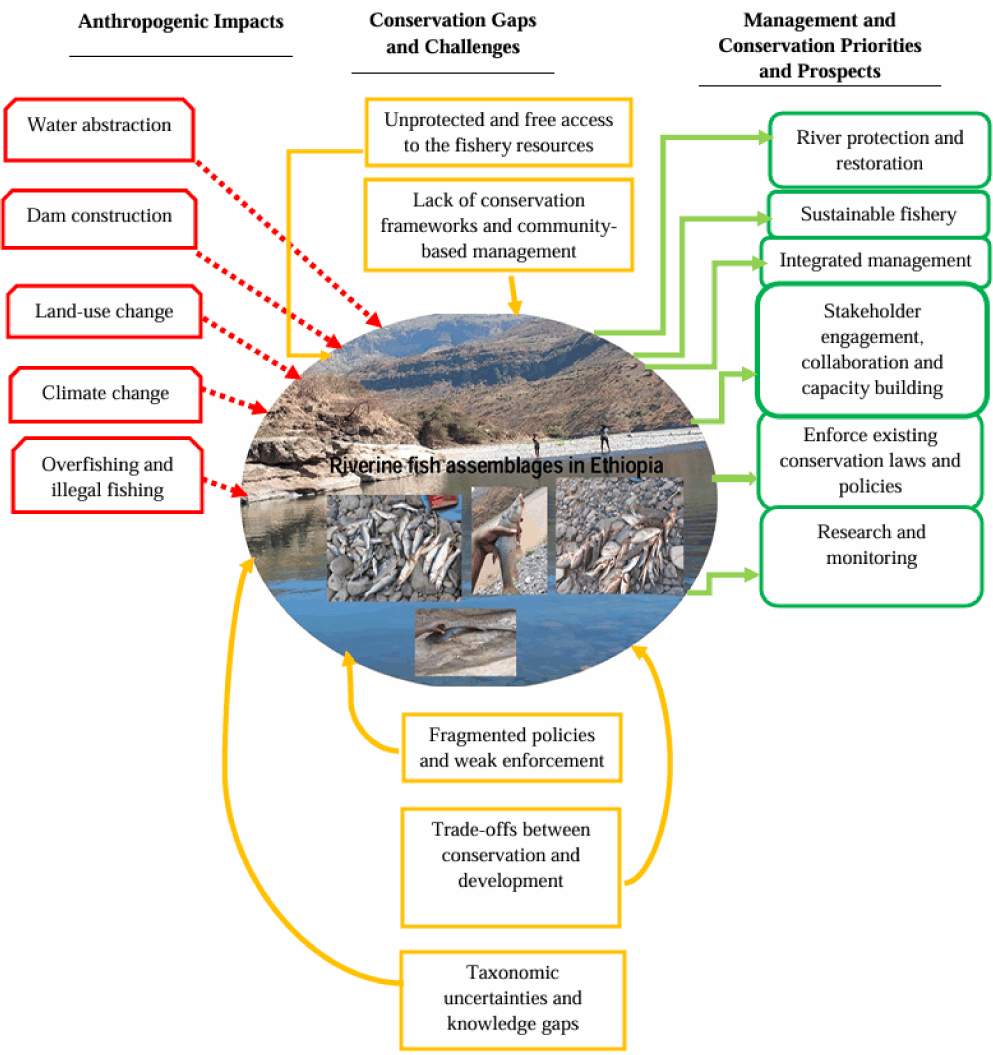
This section provides an overview of the threats and conservation issues and potential management approaches to Ethiopian riverine fish assemblages, as illustrated in Fig. 3. Anthropogenic impacts, conservation gaps, and suggested conservation priorities are all graphically categorized in this schematic diagram. Rapid population growth and economic development in Ethiopia degrade habitats and change riverine systems, imposing numerous anthropogenic stressors on river fish assemblages (Wendimu et al., 2024). The schematic diagram highlights major stressors such as water abstraction, dam construction, land-use changes, climate change, and overfishing, all of which disrupt river ecosystem and fish population. Effective fish conservation is further hampered by a lack of conservation frameworks, fragmented policies, and fish taxonomic uncertainties. To alleviate these threats and conservation problems, management strategies like river restoration, sustainable fishing methods, stakeholder involvement, and the enforcement of conservation laws are prioritized.
Ethiopian lakes and rivers are also experiencing increasing human use pressure from irrigation, household water supply and industry (Lemma & Desta, 2016). Hydroelectric and irrigation projects in Ethiopia result in lower water levels, altered pH, and altered nutrient and sediment levels, which impact fish breeding grounds and reduce fish stocks (Vijverberg et al., 2012).
Agriculture in Lake Tana and tributaries extensively uses water resources affecting Laboebarbus spawning sites (Gebremedhin et al., 2013). Water abstraction for irrigation in April–May (extreme dry months in Ethiopia) disconnects tributary pools and affects Labeobarbus juveniles’ downstream migration (Anteneh et al., 2013). The Gumara River, a cardinal tributary of Lake Tana, has hydrological changes for 20 years because of unregulated pump irrigation and poor catchment management (Abebe et al., 2021). The abundance of juvenile Labeobarbus declined in Gumara River pools after the lake disconnection due to excessive irrigation abstraction (Anteneh et al., 2013, Zelalem et al., 2022).
Excessive withdrawal of water from the Awash River for agriculture, industry and domestic use degrades habitats, reduces water availability and changes flow patterns, which affect aquatic ecosystems, riparian areas and species and leads to a decline in biodiversity (Water and Land Resource Centre, 2024). According to Teshome’s 2019 study, irrigation during dry seasons in the Afar region’s Tendaho reservoirs reduces surface area and depth, and results in a significant loss of fish yield in years with high evaporation, low rainfall and excessive irrigation.
In Ethiopia, irrigation schemes have been implemented by diverting and damming rivers in different parts of the country (Mequanent et al., 2022). Several authors examined the effects of large weirs and dams on fish abundance and diversity in Ethiopia (Anteneh et al., 2013; Gebremedhin et al., 2017; Mequanent et al., 2022). Large dams alter natural flow, fragment habitats and inhibit fish migration, which reduces fish populations (Gebremedhin et al., 2017; Mequanent et al., 2022).
Dams affect most migratory fishes and reduce reproduction (Gebremedhin et al., 2018; Mequanent & Mingist, 2019; Vijverberg et al., 2012). Mega hydropower and irrigation dams are under construction in the tributaries of Lake Tana (Dejen et al., 2017) and these will further restrict migratory spawning of Labeobarbus species (Anteneh et al., 2013). The Ribb dam and weir in Gilgel Abay (Mequanent et al., 2022) and Shini and Gelda rivers (Gebremedhin et al., 2017) restrict Labeobarbus spawning routes with irrigation. Gebru et al. (2019) confirmed that expansion of hydropower or irrigation dams affects water quality and quantity, and fish migration. These dams reduce downstream water flow and thus inundation of African catfish and Nile tilapia spawning areas.
Ethiopia experiences major land-use change, deforestation, settlement, and cropland expansion (Engdaw et al., 2024), leading to deterioration of water quality and biodiversity loss in freshwater bodies (Fetahi, 2019). Land use change is a threat to freshwater fish species diversity in Ethiopia (Fashing et al., 2022). Rivers are modified for agriculture affecting fisheries and livelihoods (Eyayu & Getahun, 2022a). Habitat degradation, such as changing breeding grounds or diverting rivers from lakes, causes decline in Labeobarbus stock in Lake Tana (Gebremedhin et al., 2012). Intensive farming, deforestation, overgrazing, irrigation and pollution degrade Ethiopia’s environment, decreasing fish fauna and biodiversity in different drainage basins. Englmaier et al. (2020) reported low fish species diversity in the Awash River catchment attributed to deforestation, invasive plantations, intensive agriculture, overgrazing, high nutrient input, erosion and water pollution, resulting in habitat degradation, loss and decline of fish species (Water and Land Resource Centre, 2024).
Agriculture and industrial sewage pollute Ethiopian fisheries and water bodies (FAO, 2014). Textile industries in Hawassa, Arba Minch and Koka Reservoir discharge pollutants and nutrients that affect lake and river biodiversity (Wendimu et al., 2024). Poor waste management and chemical inputs pollute rivers and wetlands, harming fish and aquatic life (EWNHS & WI, 2018; Water and Land Resource Centre, 2024). Agricultural runoff introduces nutrients and pesticides that degrade water quality. Fish decline in the Gumara River is attributed to anthropogenic pollution from towns and agricultural lands that reduces fish diversity and riparian vegetation (Abebe et al., 2021; Zelalem et al., 2022).
Ethiopia is a highly vulnerable country to climate change because of its large population and history of recurrent food insecurity (Evangelista et al., 2013). Climate change impacts include increased temperatures, changes in rainfall patterns and extreme weather which in turn impacts ecosystems and species distribution (Water and Land Resource Centre, 2024). Anthropogenic climate change in Ethiopia impacts aquatic ecosystems and the human societies that depend on them (Asmare et al., 2016; Mohammed & Uraguchi, 2013; Kebede & Gubale, 2016). Vibration of rainfall leads to occasional high runoff and increase in sediment load in water bodies (Wendimu et al., 2024). Changes in temperature and precipitation affect rivers (Abebe & Kebede, 2017).
Climate change is affecting fisheries in Ethiopia (Kebede & Gubale, 2016). Longer droughts reduce fish habitats (Gebru et al., 2019; Mequanent et al., 2022; Teshome, 2019). Higher temperatures increase evaporation, decrease lake inflow and lower reservoir levels. Littoral areas, where juvenile fish breed, are affected, decreasing new fish recruitment and annual yield (Lakew et al., 2016). Precipitation and seasonal variability are driving inland fisheries shifts (Asmare et al., 2016). Warmer water temperatures reduce fish stocks by changing trophic status and water quality (Teshome, 2019; Wendimu et al., 2024). Lower water levels increase pollutant and nutrient concentrations, threatening biodiversity and productivity (Lakew et al., 2016).
Climate change impacts fishing resources and fish output for human consumption (Tewabe, 2015), making fishing households more vulnerable; thus, fishers catch whatever fish they can regardless of size (Lakew et al., 2016; Temesgen, 2017). Climate-driven changes in temperature and flow regimes are likely to affect key biological processes, including recruitment success, migration timing, and spawning cues. Reduced flood magnitude and duration may limit access to spawning and nursery habitats for potamodromous species (e.g., Labeobarbus spp.), while increasing water temperatures may exceed thermal tolerances for early life stages, reducing juvenile survival and long-term population resilience.
Overfishing occurs in Ethiopia due to weak legal framework, unsuitable fishery law enforcement and unregulated activities (Wendimu et al., 2024). Overfishing depletes Omo River and Tekeze Basin fish stocks (Water and Land Resource Centre, 2024). Overfishing decreases fish stocks and affects aquatic ecosystems (EWNHS & WI, 2018). Fish production in Lake Tana decreases (Dejen et al., 2017), 93% fishers attributed lower Omo River overfishing (Wakjira, 2016). Overfishing decreases L. intermedius and reduces fish availability in local markets (Dadebo et al., 2013).
Overexploitation and resource depletion are results of fishing gear types and mesh sizes. In Ethiopia most gears are traditional and smaller than recommended (Brook, 2012; Deng, 2020; Kebede & Gubale, 2016). Gillnets, beach seines, long lines, hook-and-line and cast nets are used. Scoop nets and wire-plant baskets are used in rivers. Lake Ziway research showed 43.33% overexploitation due to small mesh sizes (Muluye et al., 2016). Lake Tana fishers use undersized (5–7 cm) stretched monofilament gillnets (Tewabe, 2015), which are imported from Sudan since 2008 and causing fish devastation in Lake Tana (Gebremedhin et al., 2013).
Fishers are not aware of the timing of first sexual maturity leading to overfishing (Wendimu et al., 2024). Muluye et al. (2016) reported that 50.6% of fishers were not aware whether fish captured were mature or immature. Unsustainable fishing practices with destructive gears and targeting breeding populations have resulted in species declines and threaten biodiversity and food security (Deng, 2020). Illegal commercial gillnet fishing in Lake Tana’s wetlands and river mouths (Gebremedhin, 2014; Gebremedhin et al., 2012) and in Alitash National Park’s floodplain rivers (Eyayu & Getahun, 2022a) has reduced fish abundance. Poisons from different plants are applied in diverse ways in different waterbodies. Asmare et al. (2016) reported poisoning with Millettia ferruginea seeds and Balanites aegyptiaca bark in Wonchit and Jemma Rivers. Poisoning waterbodies is illegal and causes damage to aquatic ecosystems.
Fish and fishery management faces several challenges in Ethiopia, especially in rivers. We intend to assess key conservation gaps and challenges as follows.
Capture fisheries are often considered as common resources, so overexploitation is difficult to avoid. Many fish stocks are not owned by fishers’ cooperatives, so led sustainable use fisheries difficult. Lack of stewardship asset towards communal pool resources often experience lacks responsibility for resource protection (Lakew et al., 2016). This leads to overfishing, fish stock depletion, and ecosystem and livelihood damage.
Ethiopia has published limited studies on fish species (Getahun, 2017); most studies are fragmented and mainly academic in nature limiting practical fisheries development and management (FAO, 2012). Due to a lack of thorough inventories and the need for taxonomic revision for numerous groups, estimates of fish diversity in Ethiopian inland water bodies are still imprecise (Anteneh et al., 2023). Every year, new species are discovered, even in the most studied hydrographic basins (Wakjira & Getahun, 2017). Scientific information mainly covers Lake Fishery that is limited in coverage and seasonality. River fish assemblages in major basins are minimally studied (Eyayu & Getahun, 2024; Getahun, 2017; Mekonen & Hailu, 2021) limiting evaluation of temporal change due to environmental pressures or human activities. Basic biological and ecological information including maximum sustainable yield (MSY), fish biology and environmental conditions is required for effective fishery management (Lakew et al., 2016).
The Ethiopian National Fisheries Proclamation was promulgated in 2003 and provides guidelines for the conservation of the resource, food safety and aquaculture development (Vijverberg et al., 2012). Federal and regional fishery laws exist that address overfishing control, fishing gear regulation and the introduction of exotic species (Water and Land Resource Centre, 2024). Economic difficulties, ineffective administration and lack of expertise hamper fisheries development and are contributing to overexploitation through unwise fishing practices (Kebede & Gubale, 2016).
Ethiopia does not have a coherent conservation framework for rivers, and community-based fishery management is rarely implemented. Fishery co-management was developed for most lakes and reservoirs in 2013, but this does not prevent overexploitation. Low fishing community participation in fisheries management hinders implementation of policy, which is a challenge for sustainable fisheries resource management.
According to the FAO (2014), factories, agriculture and sewage are the sources of major pollutants affecting Ethiopian water bodies and fish, posing a serious threat to fisheries. The hydropower plant construction in Ethiopia illustrates the conflict between economic development and biodiversity conservation. Dams provide useful resources but also threaten the local ecosystems and fish population, making it difficult for policymakers and stakeholders to find sustainable solutions (Water and Land Resource Centre, 2024).
Fish biodiversity conservation and management are necessary to protect fish species and their habitats. Conservation means maintaining healthy populations, preserving natural habitats and sustaining fisheries. Management means actively managing fish populations and habitats by balancing ecological, social and economic factors (Meinam et al., 2023). Therfore, in response to anthropogenic impacts on Ethiopian rivers, the following multi-faceted approaches are recommended, including critical components of fish management and conservation priorities.
Fish biodiversity management involves protecting and restoring habitats, including spawning grounds, nursery areas and migration routes (Meinam et al., 2023). Protecting and restoring critical habitats (e. g., spawning and nursery areas) is critical for conserving fish assemblages. Efforts should maintain water quality and habitat connectivity to support fish migration and reproduction (Anteneh et al., 2023). Ethiopia has initiated a green legacy program focusing on reaforestation and sustainable forest management to reverse deforestation, land degradation and biodiversity loss (Lambert & Deyganto, 2023). The Ethiopia’s Climate Resilient Green Economy (ECRGE) initiative aims to restore degraded lands, conserve water resources and promote ecological balance, protecting vulnerable ecosystems and combating drought-induced desertification (Federal Democratic Republic of Ethiopia , 2015).
Sustainable fisheries management is key to preventing overfishing and promoting responsible practices (Kebede & Gubale, 2016). Improved regulatory frameworks, community outreach and enforcement are needed (Meinam et al., 2023). Fisheries management plans on Ethiopian rivers include catch limits, gear type, no-fishing zones and selective techniques (Water and Land Resource Centre, 2024). Implementing sustainable fishing practices is key to preventing overfishing and ensuring long-term viability of fish populations. This includes implementing catch limits, protecting breeding grounds and promoting selective fishing techniques to reduce bycatch (Wilson et al., 2021). Prohibiting illegal fishing and increasing public awareness are key. Strengthening monitoring and enforcement provides long-term solutions for river ecosystems and communities (Desalegn & Shitaw, 2021).
The Water and Land Resource Centre (2024) calls for an integrated approach to water resources management, with biodiversity conservation and sustainable use at the centre. Such an approach balances ecological health and human needs and requires collaboration between stakeholders such as government agencies, communities and NGOs. Managing fish assemblages in Ethiopian rivers requires integrated techniques such as watershed planning, rainwater harvesting and efficient irrigation systems (Water and Land Resource Centre, 2024).
Overall, the implementation of fishery protection legislation in Ethiopia is weak, and a number of measures are needed including strengthened enforcement, improved government coordination, increased penalties for illegal fishing, investment in technology, stakeholder collaboration and improved water quality standards (Kebede & Gubale, 2016).
Effective management of fish biodiversity requires collaboration among fishers, local communities, scientists, policymakers and conservation organizations (Meinam et al., 2023). Education programs increase biodiversity awareness and community participation in decision-making (Water and Land Resource Centre, 2024). Community-based management with local fishers is more effective and sustainable (Englmaier et al., 2020). Community-led programs, such as fisheries management and participatory forest management, improve biodiversity and fishery stewardship (Water and Land Resource Centre, 2024). Tecklie (2024) found stakeholder collaboration improves fish population resilience and habitat. Indigenous knowledge in fish management contributes to sustainability and community ownership (Meinam et al., 2023).
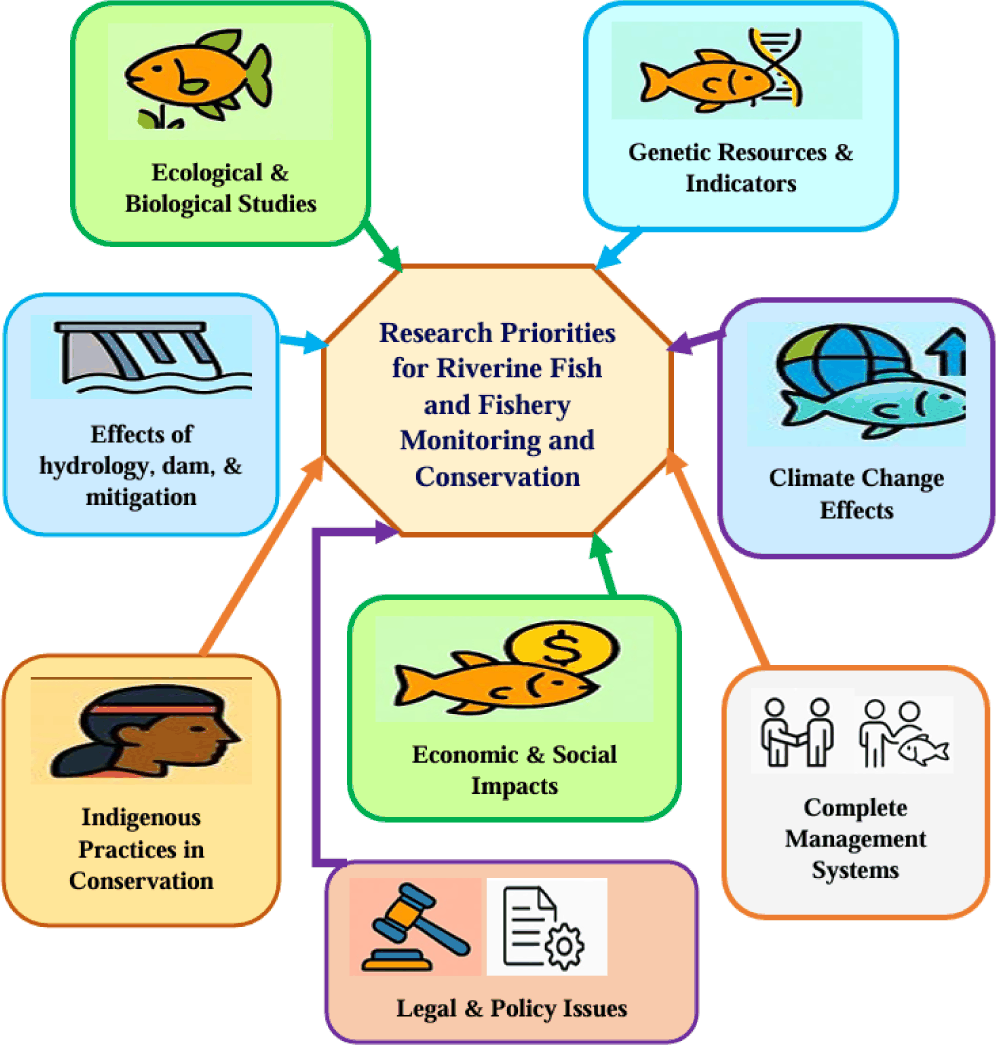
Continuous monitoring of fisheries is critical for the effective management and use of fisheries, providing information on abundance, state of the ecosystem, and human and climate impacts (Lakew et al., 2016). Exploratory fishery monitoring data are lacking for many Ethiopian rivers. A collaborative effort between research institutes and higher education institutions is required to address these issues. Such collaborative efforts enhance the understanding of fish assemblages for the conservation and management of Ethiopian rivers. Based on the above needs, the following research coverage areas should be prioritized to address these issues as summarized in Fig. 4.
-
Ecological and biological studies: The studies which next will be undertaken fill the knowledge gaps in river fish community structure, ecological roles/functional trait diversity and taxonomic diversity with environmental data/requirements thereby making a ecosystem resilient & can enforce the conservation and sustainable management of fish. Life history attributes examine any of reproductive biology; growth, habitat (population demography or) dynamics in the fish species that are relevant to conservation and sustainable fisheries. Long-term fishing strategy will give sustainable fishing, and points us therefore to a whole scale needed.
-
Studies about the effects of the environment’s hydrology, the dam, and any possible mitigation strategies: Attention should be given to water extraction, dam operations, and shifts in flow regime. A full-fledged ecological study is required to evaluate how best to balance the needs for primary power development and conservation, including examining fish passage and restoration of the habitats.
-
Genetic resources and their indicators: The investigations concerning the genetic diversity of fishes may assist in uplifting conservation efforts by focusing on sensitive species and evaluating their viability to new environmental conditions.
-
Adding or incorporating indigenous practices into contemporary conservation approaches: Local ecosystems preservation should involve the use of indigenous knowledge derived from local communities that have managed to conserve the environment properly. Such a strategy would foster more effective, sustainable, and integrated management systems by linking science and tradition (Water and Land Resource Centre, 2024).
-
Researchers should study complete management systems which unite local people and stakeholders in fisheries management alongside empowerment of indigenous communities and building strong resource capacities and ownership responsibilities.
-
The study of how climate changes affect Ethiopian biodiversity remains limited especially for fish species according to Fashing et al. (2022). Future adaptive fisheries management practices and extended sustained fishery sustainability require comprehensive research about fish species resilience and the effects of climate change on fishery diversity and productivity.
-
Fisheries researchers should analyze how fishery conservation and management influence local communities economically and socially to develop sustainable development systems.
-
Research should outline various legal issues as well as policy matters by identifying implementation challenges to help create new legislation while monitoring its effects for evidence-based decision-making. Also, evaluate current laws to propose needed improvements.
Conclusion and Recommendations
This review synthesizes current knowledge on the ecological and biological characteristics of riverine fish assemblages in Ethiopian rivers, highlighting substantial spatial variability in species composition, growth patterns, reproductive traits, and trophic ecology. Ethiopian rivers harbor diverse and unique fish assemblages, providing critical biodiversity and livelihood benefits. Assemblages differ according to altitude, hydrology, and habitat complexity. Threats include water abstraction, dams, land-use change, climate change, overfishing, and illegal practices.
Given the spatial heterogeneity of assemblage structure and biological traits, effective management should adopt river-specific and ecosystem-based approaches. These should integrate protection of environmental flows, maintenance of river–floodplain connectivity, seasonal fishing restrictions aligned with spawning periods and preservation of migratory corridors, and adaptive monitoring programs that explicitly account for climatic and anthropogenic pressures. Involving local communities through co-management arrangements and establishing monitoring programs are critical for ensuring long-term sustainability. Protecting and restoring habitat, sustainable fishing, integrated management and engaging stakeholders are among the many approaches needed. Continuous research and monitoring fill knowledge gaps that are crucial for adaptive management practices ensuring the viability of fish populations over the long term and the health of aquatic ecosystems in Ethiopia.