





Background
Type 2 diabetes is the complex metabolic disorders characterized by an abnormal glucose metabolism arising from pancreatic β-cell dysfunction and glucose intolerance (Hamza et al. 2010, Guilherme et al. 2008, Bell et al. 2001). Glucose metabolism is an essential target to treat type 2 diabetes (Rines et al. 2016, Tsuneki et al. 2004). Controlling blood glucose levels requires the establishment of balance between glucose utilization, glucose production, and glucose transport (Giugliano et al. 2008). Previous studies have emphasized adults in a state of glucose intolerance and obesity of children and adolescents are at increased risk of disrupted glucose metabolism (Wiegand et al. 2005). Furthermore, it has been established glucose intolerance and β-cell dysfunction are characteristics of the pathogenesis of impaired glucose metabolism. Hyperglycemia and glucose intolerance are suppressed by improved glucose transport, which is regulated by the GLUT transporter family (e.g., GLUT2 and GLUT4), and members of this family are essential for the controlling blood glucose level (Nishiumi et al. 2010, Yamashita et al. 2012). Herman and Kahn reported glucose transport occurs in the pancreatic β-cells, skeletal muscle, adipose tissue, and brain and that it is essentially required for blood glucose homeostasis (Herman et al. 2006).
The glucose transporter 2 (GLUT2) in pancreatic β-cells is demanded for glucose-stimulated glucose tolerance and insulin secretion, which are both reduced in diabetic mouse models (Bonny et al. 1997). On the other hand, glucose transporter 4 (GLUT4) in the skeletal muscle is a key glucose transporter and plays an essential role in the extracellular glucose transport into insulin-sensitive cells (Chang et al. 2004, Shan et al. 2011).
Seaweeds are valuable sources containing diverse bioactive substances with potentials as nutraceutical and pharmaceutical agents (Lee et al. 2010, Kiuru et al. 2014), and recent studies have reported seaweeds and their active substances can improve glucose metabolism by stimulating glucose transport in diabetes (Murugan et al. 2015, Sharifuddin et al. 2015). Ishige okamurae (IO), as an edible brown seaweed, has been shown to contain biologically active substances like diphlorethohydroxycarmalol (DPHC), ishophloroglucin A, and fucoxanthin and associated secondary metabolites (Sanjeewa et al. 2017). Furthermore, it was shown in a previous study that an ethanolic IO extract (IOE) regulated blood glucose level by increasing the level of glucose-metabolizing enzyme in the liver and promoting insulin resistance in db/db mice which is a leptin receptor-deficient model (Min et al. 2011). However, the effect of IOE has not been investigated on the glucose metabolism in the pancreas and muscle of high-fat diet (HFD)-fed mice (a well-established model of glucose intolerance) (Honors et al. 2012, Riant et al. 2009). Accordingly, we evaluated the effects of an IOE on the protein levels of glucose GLUT2 and GLUT4 and on those of linked transcription factors of the pancreas and skeletal muscle and on blood glucose levels in HFD mice.
Materials and methods
The 50% of ethanol extract of Ishige okamurae (IOE) was supplied by Shinwoo Co. Ltd. (Lot No. SW9E29SA, Republic of Korea) as previously described by Yang et al. (2019). Depending on a previous method (Ryu et al. 2018), the IOE used to this study was standardized by assuming diphlorethohydroxycarmalol (DPHC, 2.37%) via HPLC analysis. The HPLC chromatogram of DPHC is shown in Figure A1. The chromatographic analyses were carried out by an Alliance 2695 Separations Module equipped with a 2998 PDA detector (all from Waters, Milford, MA, USA) and an Agilent poroshell 120 EC-C18 column (4.6 × 100 mm, 4 μm). And, the mobile phase contained (A) 0.1% formic acid in water and (B) ACN with 0.1% formic acid. The eluting conditions of HPLC were as follows: 20–40% B for 25 min, following that re-equilibration time of the column for 10 mins. The rate of flow was kept at 0.3 mL/min and the injection volume was 10 μL. The content of DPHC was validated at Korea Health Supplement Institute (KHSI; Seongnam-si, Gyeonggi-do, Republic of Korea). The presence of DPHC was inspected by quadrupole time-of-flight lipid chromatography-mass spectrometry (Q-TOF LC-MS/MS) using an electrospray ionization (ESI) source (maXis-HD, Bruker Daltonics, Breman, Germany) at the Korea Basic Science Institute (KBSI; Ochang, South Korea).
C57BL/6N male mouse (age, 7 weeks) was purchased from Orient Bio Inc. (Republic of Korea) and housed individually in stainless steel cages at 24 °C and 45–50% RH with a 12-h light-dark cycle. Before the experimental period, all mice were fed a control chow diet for 1 week. All mice were randomly allocated to five groups (n = 3 for each group): mice were fed a chow diet for 8 weeks with 0.9% saline (wt/wt on chow diet) for the last 4 weeks (the NFD/saline group); mice were fed a 45% high-fat diet (HFD) (Research Diet Inc., USA) for 8 weeks with 0.9% saline (wt/wt on HFD) for the last 4 weeks (the HFD/saline group); and mice were fed the 45% HFD for 8 weeks with 25 mg/kg (HFD/25 IOE), 50 mg/kg (HFD/50 IOE), or 75 mg/kg (HFD/75/IOE) of IOE during the latter 4 weeks. Mice were sacrificed on experimental day 57 in accordance with guidelines signed by the Animal Care and Use Committee of Gachon University (IACUC; LCDI-2018-0112).
The intraperitoneal injection of glucose (2 g/kg of body weight) in mice that had been fasted for 8 h and blood was obtained from a tail vein 30, 60, 90, or 120 min later, as previously described (Konrad et al. 2002). Serum glucose level was measured using a blood glucose meter (AccuChek Advantage; Roche Diagnostics Corp., USA).
Paraffin blocks of pancreatic tissue were sectioned at 5 μm, laid on coating slides, dried at 40 °C for 12 h, and deparaffinized using xylene. And then, the deparaffinized tissue sections were incubated in 0.3% hydrogen peroxide (Sigma-Aldrich, USA) for 30 min, washed 3 times with PBS (phosphate-buffered saline), blocked with normal animal serum, incubated with anti-PCNA antibody (Abcam, USA), and rinsed with PBS 3 times. The tissue sections were incubated with biotinylated secondary antibodies in the ABC kit (Vector Laboratories, USA), washed 3 times with PBS, treated with DAB (3, 3-diaminobenzidine) substrate to 0.03% hydrogen peroxide for 10 min, mounted using a xylene-based DPX mounting solution (Sigma-Aldrich, USA), and photographed under the microscope (Olympus Optical Co., Japan).
The pancreas and skeletal muscle (soleus and gastrocnemius muscle) paraffin block section (5 μm) was de-paraffinized, treated with animal serums for blocking antibody binding, and incubated with anti-GLUT2 and anti-GLUT4 antibodies (Santa Cruz Biotechnology, Inc., USA) at 4 °C for 1 day. And then, the tissue sections were washed with PBS, incubated with fluorescence conjugated secondary antibody at room temperature for 1 h, stained with DAPI (4′ 6-diamidino-2-phenylindole; Sigma-Aldrich, USA) solution for 1 min, rinsed with PBS 3 times, and mounted using Vectorshield solution (Vector Laboratories). Fluorescences were detected using confocal microscopy (LSM 700; Carl Zeiss Ltd., Germany).
Total RNAs of mouse’s pancreases and skeletal muscles (soleus and gastrocnemius) were lyzed using the RNAiso Plus kit (TAKARA, Japan). Briefly, cell pellets were suspended using 1000 μl RNAiso Plus by pipetting, and lysates were mixed with 100 μl chloroform (Amresco LLC, USA) and centrifuged at 12,000×g at 4 °C for 15 mins. Obtained supernatants were mixed with 250 μl isopropanol and centrifuged, and the RNA pellets so obtained were either rinsed with 70% ethyl alcohol and centrifuged at 7500×g at room temperature for 5 min or dissolved into 30 μl of diethyl pyrocarbonate (DEPC)-treated water. The levels of RNA were quantified by a Nanodrop 2000 (Thermo Fisher Scientific, Inc., USA). In order to conduct quantitative real-time polymerase chain reaction (qRT-PCR), complementary DNA (cDNA) synthesis was performed using a cDNA synthesis kit (TAKARA). Subsequently, cDNAs, SYBR green (TAKARA), and adaptive primers were mixed and amplified in PCR (Bio-Rad, USA) to profile gene expression. Validated genes are listed in Table A1.
Paraffin block slides of pancreas tissues were deparaffinized, stained with Mayer’s hematoxylin solution (DAKO, England) for 30 s and eosin Y solution (Sigma-Aldrich, USA) for 10 s, rinsed 3 times with distilled water, and mounted using xylene-based DPX mounting solution. H&E stained tissue slide images were captured by light microscopy and pancreatic islet sizes were calculated using the Image J software (NIH, USA).
The significances of intergroup differences were identified using the Kruskal-Wallis test. And, post-hoc comparisons were determined using Mann-Whitney U test. The analysis was conducted using SPSS software, and results are expressed to means ± SDs. Statistical analysis was admitted by p values of < 0.05. An asterisk indicates significantly different from the NFD/saline group and a number sign significantly different from the HFD/saline group.
Results
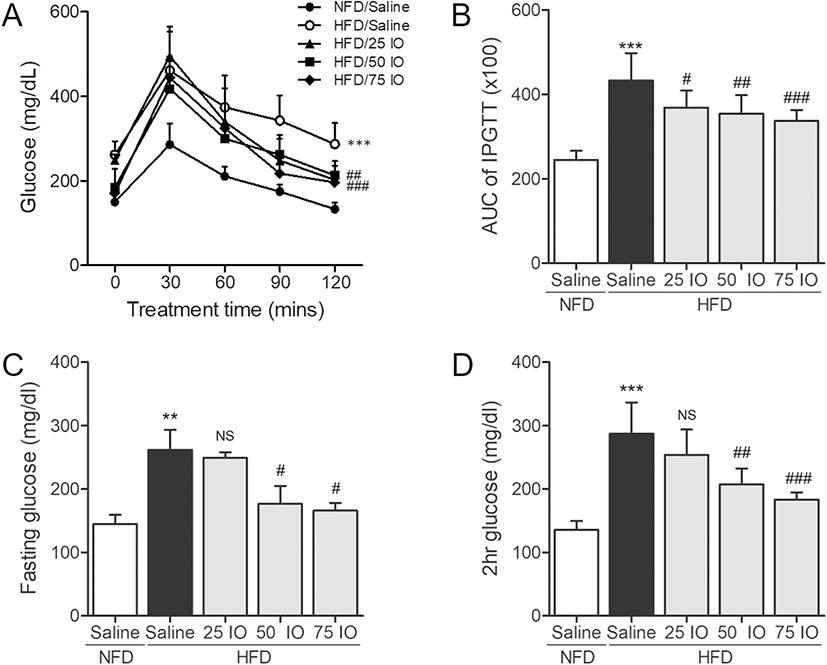
To determine whether IOE reduces elevated blood glucose level in HFD mice, we investigated glucose tolerance in HFD mice. We supplemented HFD mice feed with IOE (25, 50, or 75 mg/kg, the HFD/IOE groups) for 4 weeks after mice had been fed HFD for 4 weeks. Intraperitoneal glucose tolerance test (2 g/kg, IPGTT) was conducted at 30, 60, 90, or 120 min after injection of glucose to investigate glucose tolerance. After 8 weeks, the HFD group gained more body weight compared with the NFD group. However, the HFD/IO (25, 50, and 75 mg/kg) group showed a slightly decreased in body weight, but no significance compared with the HFD group. Body weight is shown in Figure 2A. As shown in Fig. 1a, HFD/saline controls (fed an HFD diet for 4 weeks and followed by saline for another 4 weeks) had lower glucose tolerance with higher blood glucose levels at each time after glucose injection than the NFD controls (fed a chow diet for 8 weeks). However, HFD/IOE groups (fed a HFD diet for 8 weeks and IOE at 25, 50, or 75 mg/kg for the latter 4 weeks) showed significant reductions in glucose levels at 120 min after glucose injection than were observed in the HFD group. AUC (area under the curve) levels of IPGTT were expressed to estimate the level of glucose tolerance impairment in each group. In line with the results shown in Fig. 1, the AUC of IPGTT in the HFD group was remarkably greater (p < 0.001) than in the NFD group (Fig. 1b). HFD/IOE (25, 50, or 75 mg/kg) groups showed dose-dependent decreases in the AUC of IPGTT. The maintenance of fasting and 2 h glucose levels is a major therapeutic goal in diabetes (Ratner 2001). In Fig. 1c and d, both fasting and 2 h glucose levels in the HFD group were significantly higher than in the NFD group. However, fasting and 2 h glucose levels in the HFD/IOE 50- and 75-mg/kg groups were significantly lower than in the HFD group. These results demonstrate that IOE can ameliorate glucose tolerance in HFD mice. Furthermore, we examined the improvement of IOE on glucose metabolism in the pancreas and muscle tissue of HFD mice in controlling blood glucose levels.
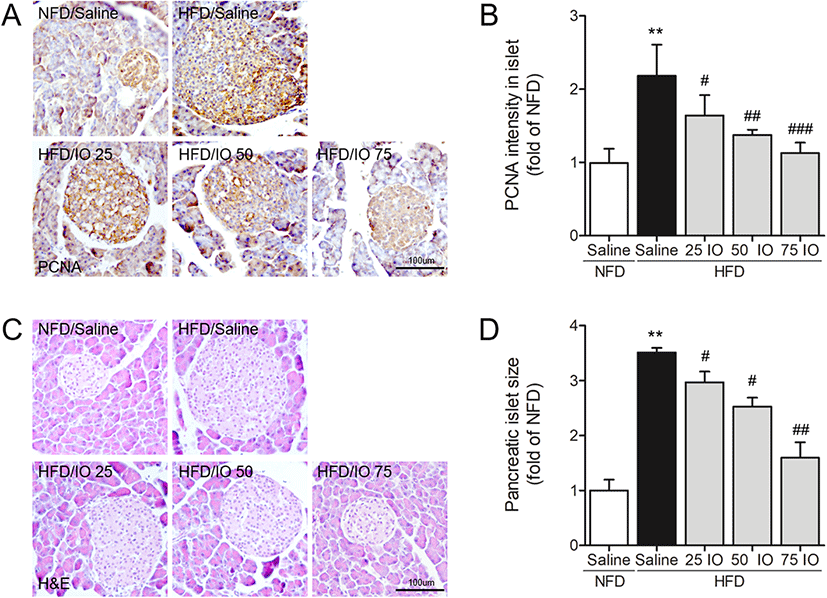
In order to investigate the effect of IOE on the morphologies and areas of the pancreatic islets of HFD mice, we conducted hematoxylin and eosin (H&E) staining and immunohistochemistry (Matveyenko et al. 2009, Sun et al. 2016). The proliferating cell nuclear antigen (PCNA, a proliferation marker) was used to detect the proliferation of β-cells (Bringhenti et al. 2013). In order to identify the effect of IOE on the proliferation of β-cells in pancreatic islets, we measured PCNA intensities (Fig. 2a, b). PCNA intensities on β-cells in pancreatic islets of the HFD group were remarkably greater than in the NFD group, whereas PCNA intensities were dose-dependently decreased in the HFD/IOE groups as compared to those of the HFD group. H&E staining pancreatic islets were larger in HFD than in NFD mice (Fig. 2c, d), and that this increase in size was significantly and dose-dependently diminished in the HFD/IOE groups (25, 50, or 75 mg/kg). These data indicated that IOE can improve β-cell function in the pancreatic islet of HFD mice.
HFD-induced diabetes is related to failure of β-cell function (Matveyenko et al. 2009, Cerf. 2006), and GLUT2 is essential for β-cell glucose sensing, which leads to glucose-induced insulin secretion (Cerf. 2006, Folli et al. 2011). In order to examine the effect of IOE on GLUT2 stimulation in pancreases of HFD mice, we evaluate the intensity of GLUT2 by immunofluorescence (Fig. 3a, b). The immunofluorescence intensity of GLUT2 in HFD mice was remarkably lower than in NFD mice (p < 0.05), and this reduction in immunofluorescence was slightly reduced in the HFD/IOE 20- and 50-mg/kg groups, but significantly reduced in the 75-mg/kg group.
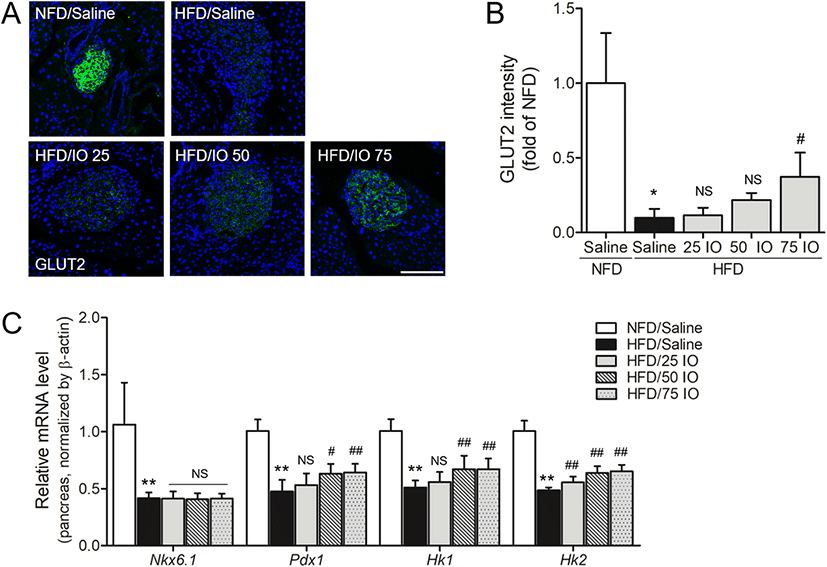
In addition, the GLUT2 transcription is controlled by pancreatic duodenal homeobox 1 (PDX1) and Hexokinase isoforms 1 (HK1) and 2 (HK2), which are also included in glucose sensing and glucose metabolism. We evaluated the effect of IOE on the mRNA levels of PDX1, HK1, and HK2 in pancreas tissues of HFD mice using qRT-PCR. mRNA levels of PDX1, HK1, and HK2 were remarkably decreased in HFD controls compared with NFD controls, which showed levels of GLUT2-related transcription factors are impaired in HFD mice (Fig. 3c). However, these falls in PDX1 and HK1 mRNA levels were significantly prevented in the HFD/IOE 50- and 75-mg/kg groups. Also, HK2 mRNA levels were significantly increased in the three HFD/IOE groups compared to those of HFD controls. These results showed that the increased GLUT2 and its related transcription factors PDX1, HK1, and HK2 by IOE can improve glucose metabolism in the pancreas.
GLUT4 levels and the levels of its related transcription factors were evaluated in the skeletal muscle of HFD mouse for glucose metabolism, which is the primary site of insulin-mediated GLUT4 translocation (Shepherd et al. 1998). The GLUT4 translocation into the membrane of the skeletal muscle in HFD mice was significantly decreased compared with NFD mice (p < 0.01; Fig. 4a, b), and GLUT4 translocation was significantly greater in the HFD/IOE groups than in HFD groups (p < 0.05). These observations suggest IOE can improve glucose metabolism in HFD mice by increasing translocation of GLUT4 into the membrane of the skeletal muscle.
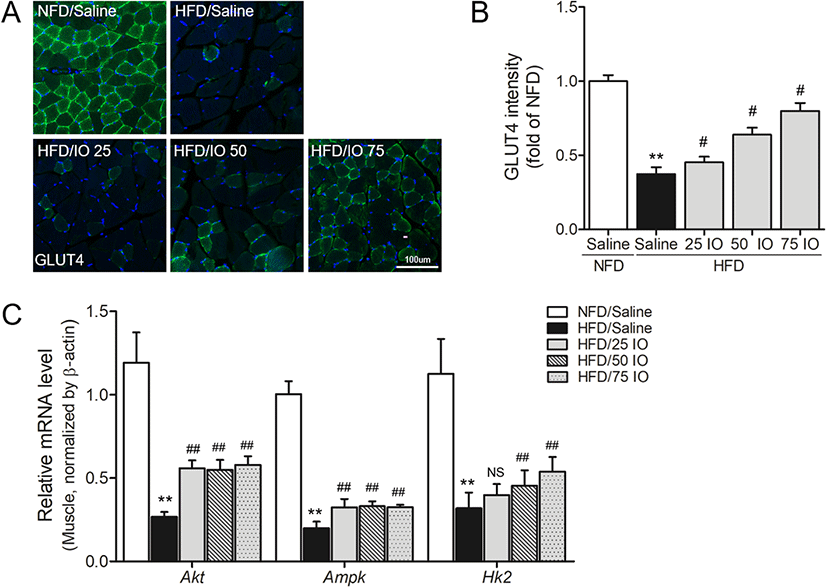
Glucose metabolism in the skeletal muscle is achieved by activation of HK2, phosphoinositide-3-kinase (PI3-K)/AKT, and AMP-activated protein kinase (AMPK) pathways (Fukumoto et al. 1988, Tsuchiya et al. 2010). We measured the mRNA level of AKT, AMPK, and HK2 in the skeletal muscle tissue of HFD mice by qRT-PCR to assess the related level of GLUT4 and glucose metabolism by IOE. Consistent with our GLUT4 translocation results, qRT-PCR showed the mRNA level of AKT, AMAPK, and HK2 in the skeletal muscle were remarkably lower in HFD than in NFD controls (Fig. 3c), but that the mRNA levels of AKT and AMPK in the three HFD/IOE groups were significantly increased compared to those of the HFD controls (p < 0.01), and HK2 mRNA levels in HFD/IOE 50- and 75-mg/kg groups were significantly higher than in HFD controls (p < 0.01). These results demonstrate that IOE enhanced the transcription of GLUT4 and those of its related transcription factors AKT, AMPK, and HK2 in our mouse HFD model.
Discussion
Given that the underlying molecular mechanisms of type 2 diabetes with insulin resistance are accompanied by β-cell dysfunction, which is probable that the cross-talk between the pancreatic cells and skeletal muscle plays an important part. Thus, identification of a novel therapeutic agent that can control the glucose metabolism by enhancing the function of cells is key to restore the blood glucose homeostasis in type 2 diabetes patients.
Of note is that IOE has been announced to have various biological activities related to diabetes. For example, it has been reported to inhibit α-glucosidase, improve glucose tolerance, upregulate levels of hepatic glucose metabolites, and reduce insulin resistance in db/db mice (Min et al. 2011, Ryu et al. 2018, Heo et al. 2009). However, the effect of IOE on functions of glucose transport in the pancreases and skeletal muscle tissues is yet to be assessed in HFD mice that have been fed with an imbalanced diet to induce mild hyperglycemia, a condition that may resemble the pre-diabetic state in humans. Therefore, we firstly confirmed that the decreased glucose transporter function in the pancreases and skeletal muscle tissues of HFD mice fed for 4 weeks was significantly ameliorated alongside improved glucose metabolism upon IOE administration for another 4 weeks.
The severe glucose intolerance in HFD mice with decreased insulin sensitivity may decelerate β-cell function in pancreatic islets, leading to excessive β-cell proliferation and increased pancreatic islet size (Matveyenko et al. 2009, Carey et al. 1996, Butler et al. 2003). Hull et al. have shown that rodents fed with an HFD to induce insulin resistance develop increased pancreatic islet size due to hyperplasia rather than hypertrophy of β-cells (Hull et al. 2005). We herein observed that the decelerated β-cell function in the pancreatic islet of HFD mice was noticeably improved by IOE administration, probably because of the restoration of the GLUT2 level. The major glucose transporter GLUT2 in hypertrophied pancreatic islets is associated with glucose sensing to stimulate insulin secretion. The mechanism of the improvement of the pancreas function by IOE through GLUT2 upregulation in the islets is linked with the regulation of PDX1, HK1, and HK2 mRNA levels, enabling acquisition of glucose-responsive insulin secretion (Frantz et al. 2013). PDX1 mutant β-cells have been shown to activate the islet-specific GLUT2 and amyloid polypeptide, and these two factors have been shown to be low in diabetic mice and patients (Nicolino et al. 2010, Suzuki et al. 2003, Waeber et al. 1996). HK1 and HK2 play key roles in glucose sensing by catalyzing glycolysis through which glucose is phosphorylated to produce glycos-6-phosphate (Hashimoto et al. 2005). Epstein et al. have reported that the elevated hexokinase activity of both HK1 and HK2 in β-cells can reduce the blood glucose level in diabetes patients (Epstein et al. 1992).
The skeletal muscle of HFD mice is known to be defective in the membrane translocation of GLUT4 alongside impaired glucose metabolism and reduced insulin-stimulated glucose transport (Fukumoto et al. 1988, Tremblay et al. 2001, Savage et al. 2007). Increased GLUT4 translocation by IOE probably facilitates glucose uptake of the skeletal muscle in HFD mice, activating the PI3K/AKT and AMPK pathways, thereby regulating the glucose metabolism, including glycogen synthesis, gluconeogenesis, and glucose transport as well as the energy expenditure (Schultze et al. 2012). Consistent with these findings, we found that the mRNA level of HK2, which promotes glucose transport and utilization, was also upregulated in HFD mice fed with IOE (Fukumoto et al. 1988).
In this study, our findings are summarized as follows: (1) IOE supplementation ameliorated blood glucose intolerance, (2) improved glucose metabolism in the pancreas, and (3) enhanced glucose transporters in the skeletal muscle. Additionally, our results suggest that the euglycemic effects of IOE in HFD mice are because of the stimulation of the glucose metabolism.
Conclusion
This study shows IOE (an ethanolic extract of Ishige okamurae) had euglycemic effects in our HFD-induced mice model of diabetes and that these effected were associated with improved glucose metabolism in the pancreas and skeletal muscle and lower blood glucose levels. We suggest that IOE is considered a potential starting point for developing the therapeutic or functional foods targeting the alleviation of diabetes.