






Introduction
Approximate 450 large fish species with about 50 commercially important fisheries resources occur in the Yellow Sea (Lee, 2004; NFRDI, 2004). Among these commercially important species, anchovy (Engraulis japonica), small yellow croaker (Larimichthys polyactis) and yellow goosefish (Lophius litulon) were caught as target species for bottom trawls, dragnets and stow nets in Korean fisheries (NFRDI, 2004, 2005). These three species distribute along the coasts of entire Yellow Sea region of Korea, and are consumed widely in Korea (NFRDI, 2004). Fishery harvests of the fish species have been fluctuated over decades in their distribution ranges (Kim et al., 1997; Lluch-Belda et al., 1989; NFRDI, 2004; UNDP/GEF, 2007a), even though they have been managed based on traditional management methods such as the closed season and closed area of fishery (MIFAFF, 2010). In Korea, ecosystem-based stock rebuilding program initiated in 2006 (NFRDI, 2006). Stomach content analysis was proposed as a method to verify trophic conditions for important fishery resources, further better understanding stock conditions of important fishery species (Zhang et al., 2009).
Analysis of stomach contents helps understand their prey composition and preference, further forecast and assess the population variations in the Yellow Sea. Stomach contents of three commercially important species, anchovy, small yellow croaker and yellow goosefish were investigated.
Anchovy is a significant food source for almost every predatory fish and birds within its distribution ranges from egg to adult stage (Hao et al., 2003; Hayasi, 1967). Small yellow croaker is widely distributed over Asia’s continental shelf in the East China Sea and the Yellow Sea (NFRDI, 2004; UNDP/GEF, 2007b, 2008), and is one of important preys of yellow goosefish (Cha et al., 1997; Choi et al., 2011). Yellow goosefish is a valuable ground fish in Korean water (Cha et al., 1997, 1998). The fish is one of the top level predators in Korean marine ecosystem with a big mouth, mostly feeding small yellow croaker and large anchovy (Cha et al., 1997, 1998).
This study focuses on documents the details of stomach contents of target species and helps fisheries researchers understand prey selectivity and composition of the target fish species.
Materials and Methods
Trawl surveys for the target fishes were conducted by the RV Tamgu-8 of the West Sea Fisheries Research Institute (WSFRI) in the Yellow Sea, Korean side Spring (May 15–21) and Fall (September 23–30) 2008 as the regional stock assessment of the United Nations Development Programme/Global Environment Facility (UNDP/GEF) Yellow Sea Large Marine Ecosystem (YSLME) survey to determine the condition of stocks and manage fisheries into the future (UNDP/GEF, 2007a). Fish were trawled at 12 stations and the hauling period was 60 minutes at each station (Fig. 1). Fish samples collected with a bottom trawl net (20 mm mesh) were classified by taxa and the body length and body weight of them were measured on board. After measured lengths and weights, fish stomachs were removed and preserved in 10% formalin. Fish stomach contents of 50 individuals each species were analyzed to species level, if possible. Stomach weights (including stomach and its contents; net stomach weight excluding the contents) were weighed, and stomach fullness and digestion condition were determined by the international scale, for stomach fullness (0–4), where 0 was empty and 4 was 100% full, for stomach fullness and digestion condition (0–5), where 0 was no digestion and 5 was empty stomach, were determined.
Results
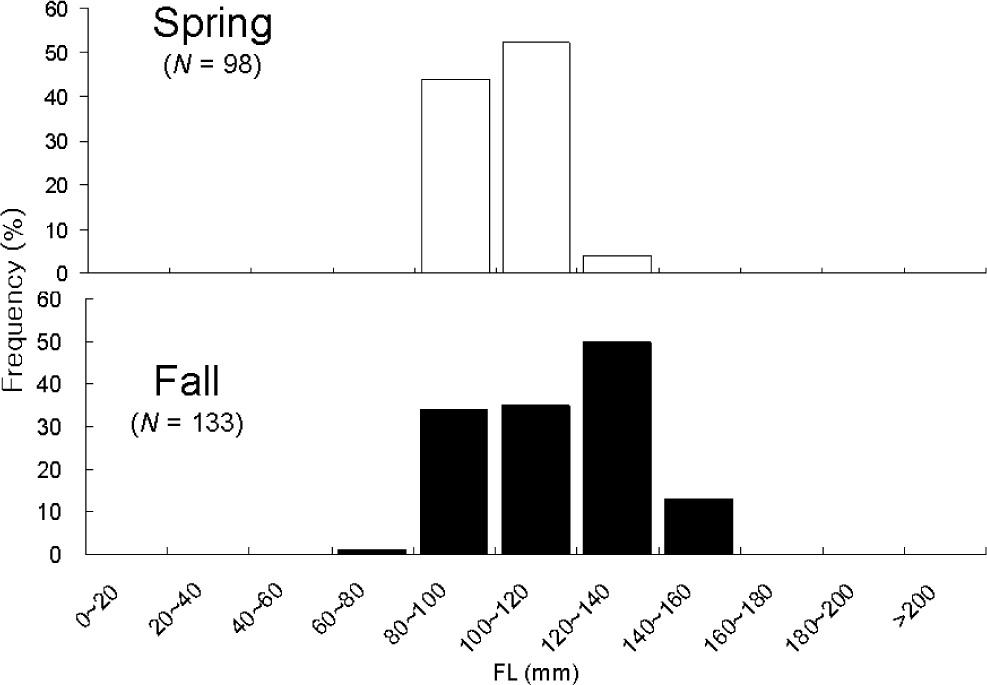
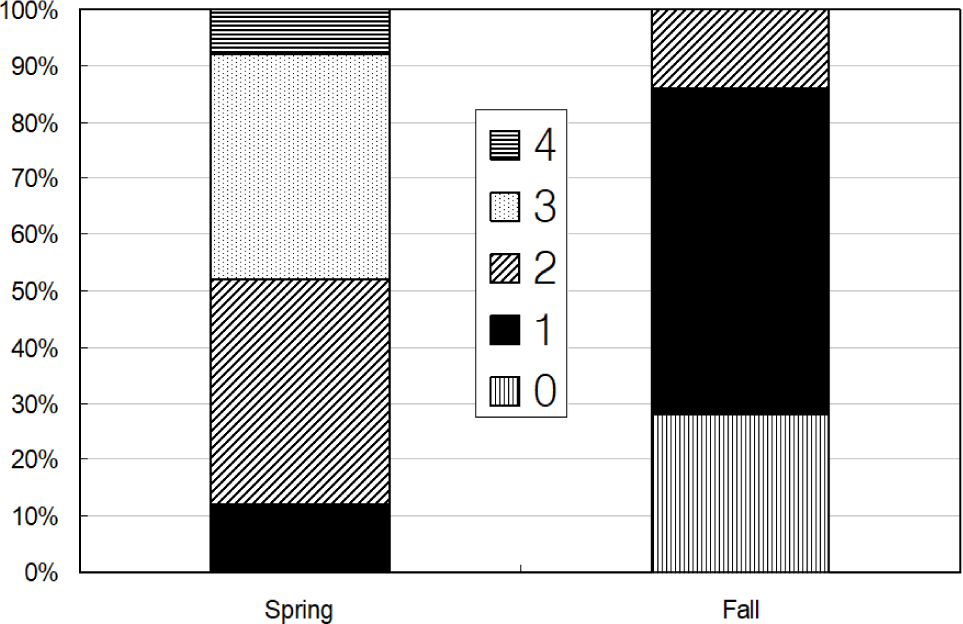
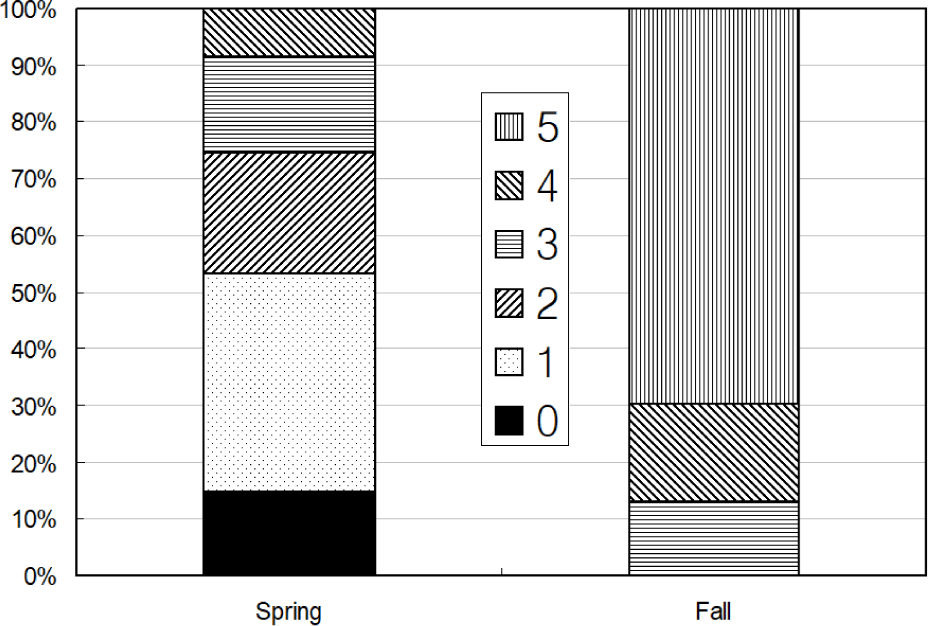
Total 231 anchovy (98 in Spring and 133 in Fall) were measured (Fig. 2). Total lengths of anchovy ranged from 60 mm to 160 mm in Spring and Fall 2008 (Fig. 2). In Spring, most anchovy size ranged within 80–120 mm while in Fall that ranged from 80 mm to 160 mm. One half of the stomachs were 25% and 50% full and the other half of stomachs were more than 50% full in Spring (Fig. 3). In Fall, most stomachs were close to empty and 14% of stomachs were 50% full. In Spring approximately one half of the stomachs had less than digestion rate of 50% and the other half of stomachs had more than digestion rate of 50% (Fig. 4). However, in Fall more than 70% of stomach contents were almost fully digested. Stomach contents consisted of 7 taxa of mysids, decapods, chaetognaths, amphipods, euphausiids, copepods and unidentified crustaceans (Fig. 5). In Spring, stomach contents were mostly occupied by copepods and euphausiids. In Fall, more than 80% of stomach contents were copepods. In Spring, proportion of copepods in the stomach contents decreased with increasing body lengths while that of euphausiids and amphipods increased with increasing body sizes (Fig. 6). In Fall, proportion of copepods in the stomach contents decreased as anchovy body size increased. Proportion of amphipods increased up to 120 mm of anchovy body size after then it decreased (Fig. 6).
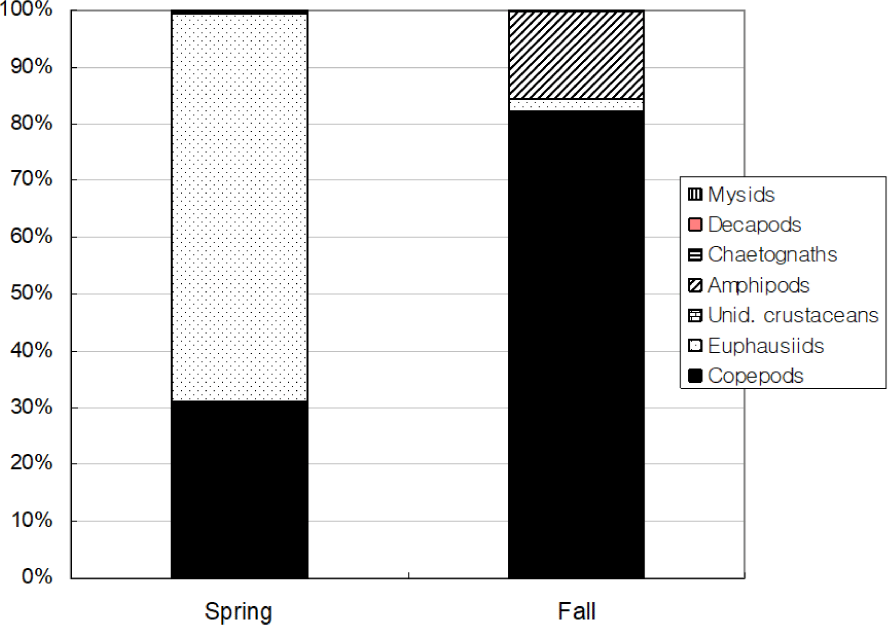
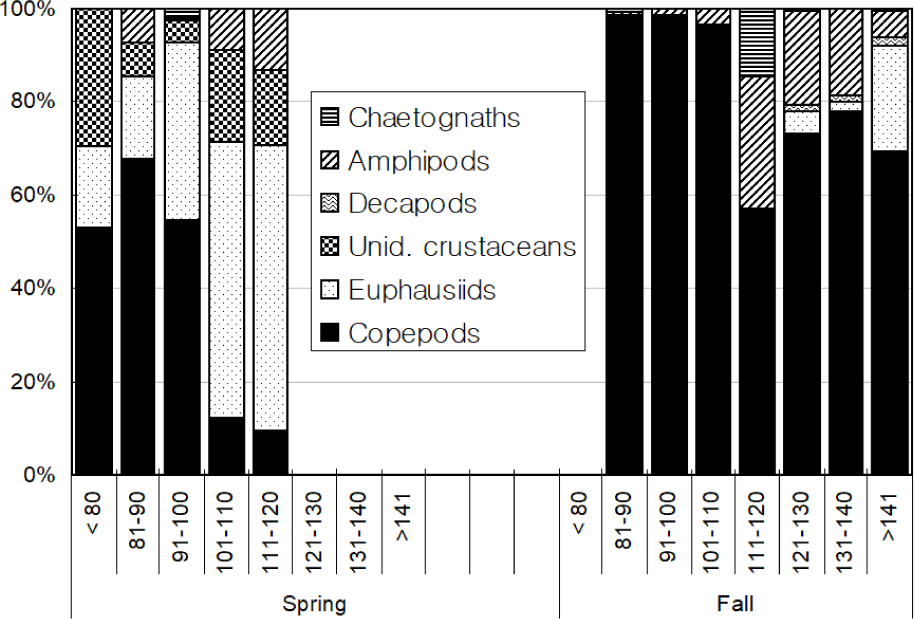
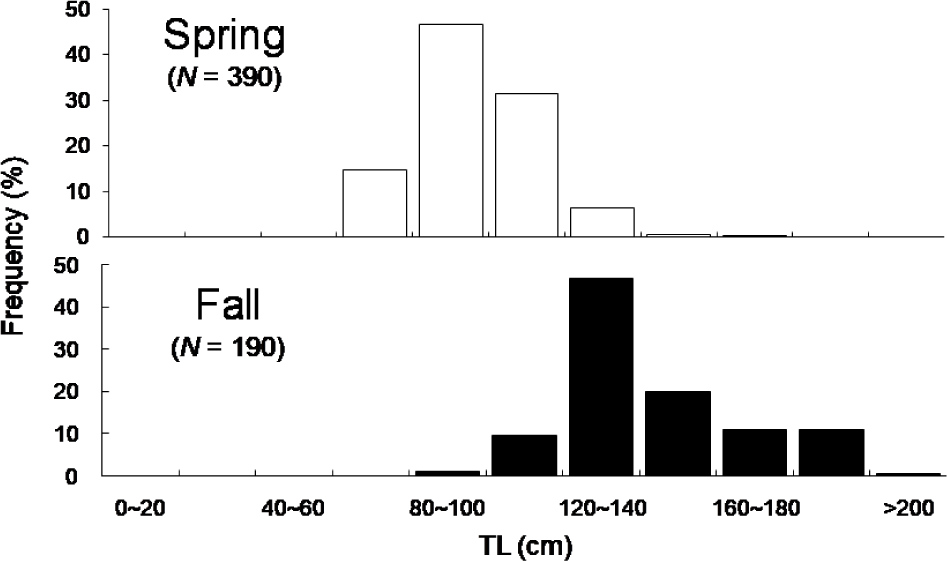
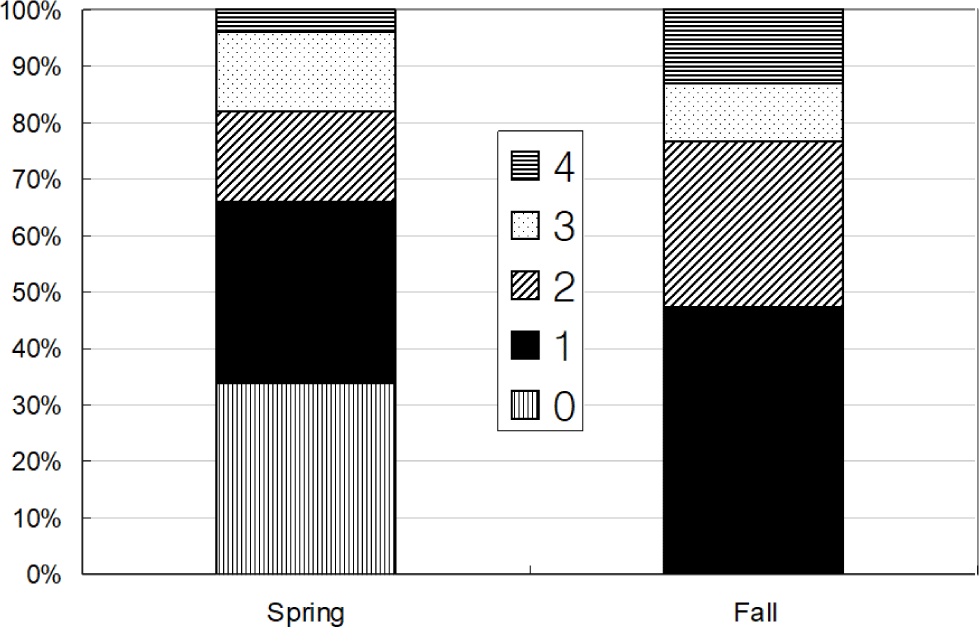
Total 580 yellow croakers (390 in Spring and 190 in Fall) were measured (Fig. 7). In Spring, the size of yellow croaker dropped in 60–140 mm while in Fall that dropped in 100–200 mm. Overall, sizes of yellow croaker shifted from smaller to larger with changing seasons. In Spring, 66 % of the stomachs were less than 25% full and 34% of stomachs were more than 50% full (Fig. 8). In Fall, 48% of stomachs were 25% full and the rest of stomachs were more than 50% full. In Spring, 73% of the stomachs had less than the digestion rate of 50% and 27% of stomachs had more than digestion rate of 50% (Fig. 9). However, in Fall more than 67% of stomach contents were almost fully digested. Stomach contents consisted of 8 taxa of molluscs, mysids, decapods, chaetognaths, amphipods, euphausiids, copepods and unidentified crustaceans (Fig. 10). In Spring, stomach contents were mostly occupied by copepods and euphausiids. In Fall, more than 73% of stomach contents were composed of euphausiids. In Spring, proportion of copepods in the stomach contents decreased with increasing body lengths while that of euphausiids and amphipods increased with increasing body sizes (Fig. 11). In Fall, no specific pattern was discovered. However, proportion of euphausiids in the stomachs was more than 50% at almost all size categories except 151–160 and 171–180 mm categories. Particularly different pattern in the anchovy stomach contents in Fall were higher proportion of decapods and molluscs in the larger anchovy compare to those in Spring (Fig. 11).
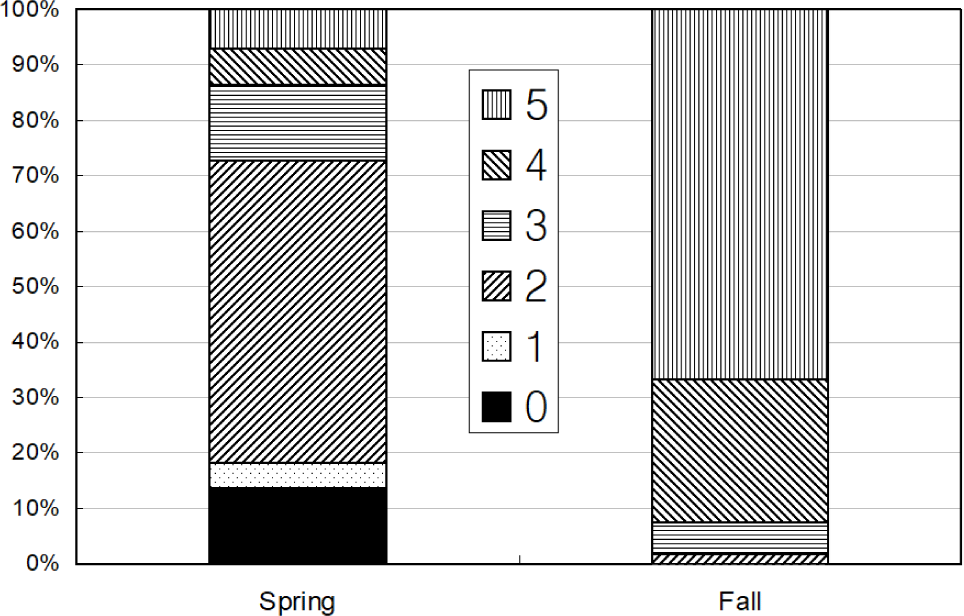
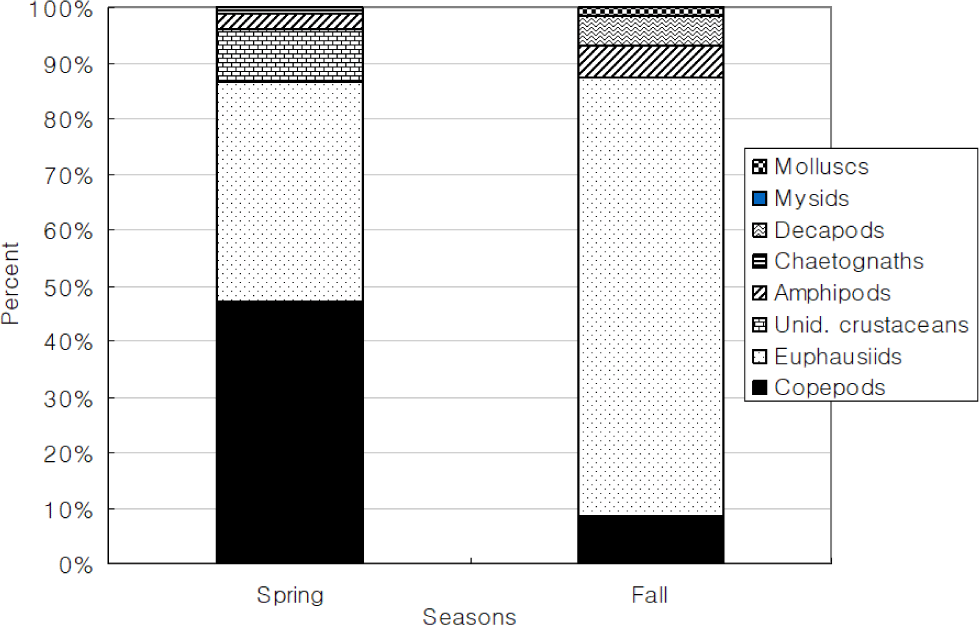
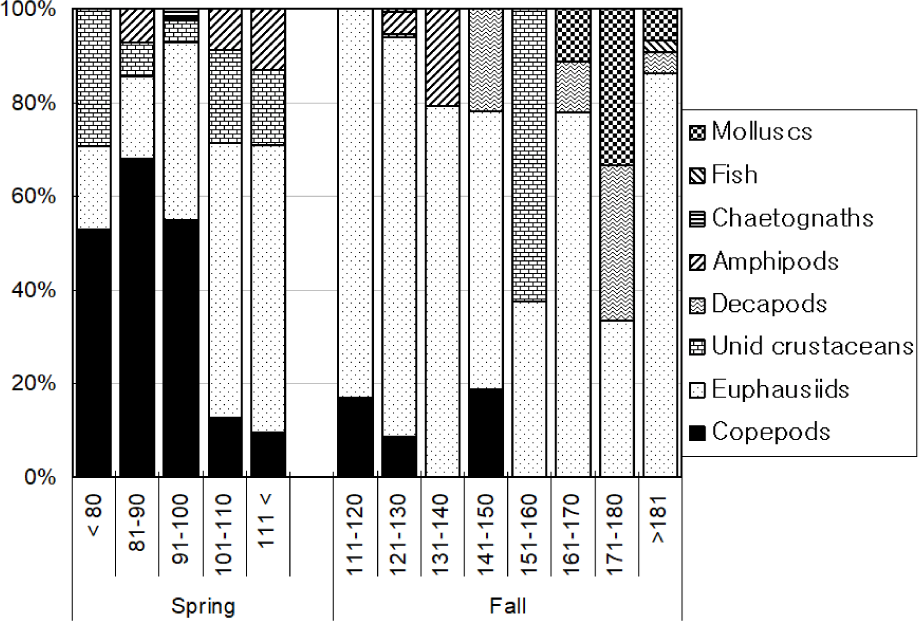
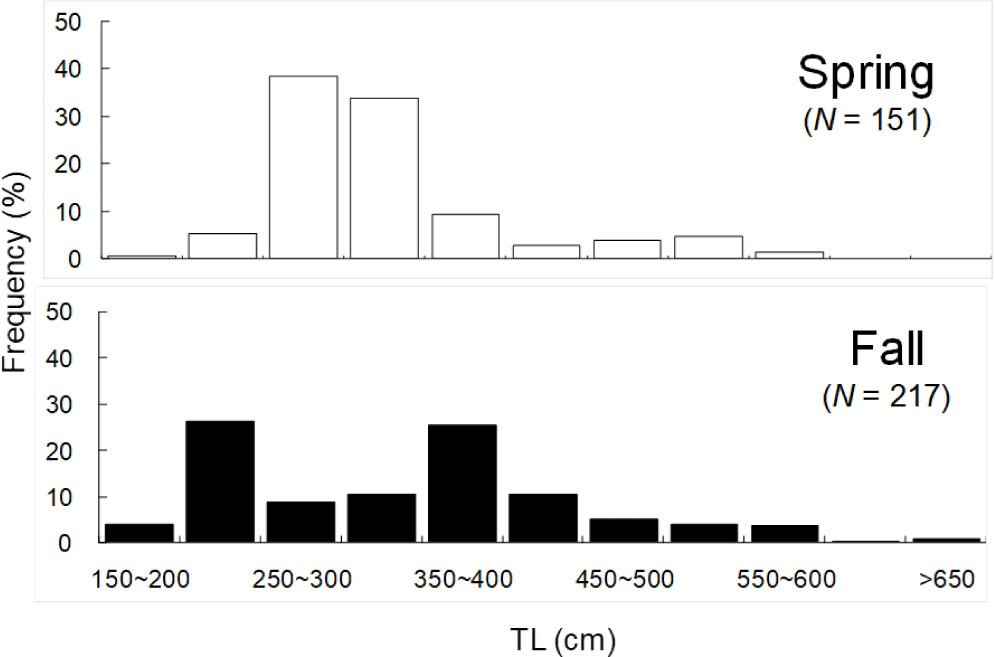
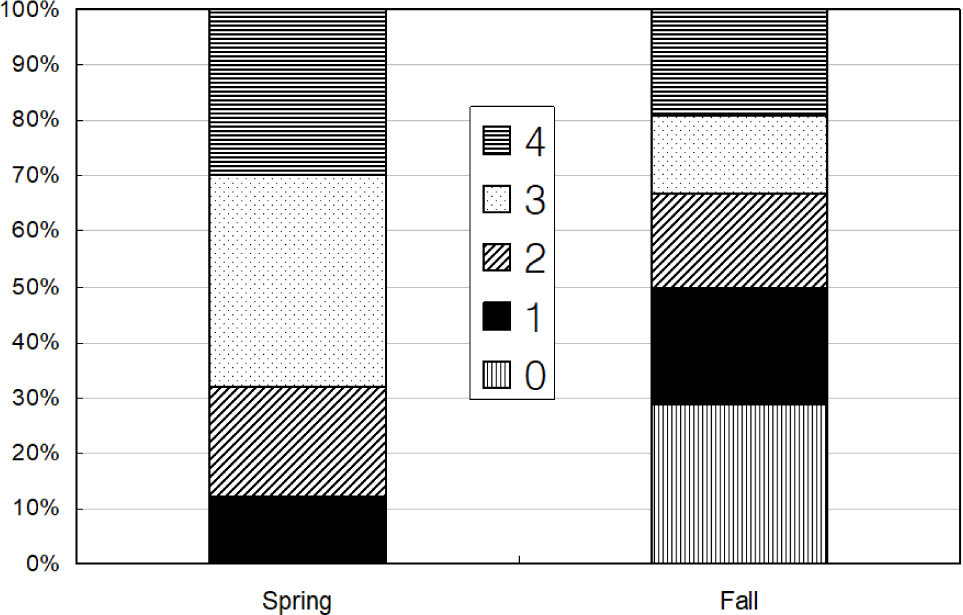
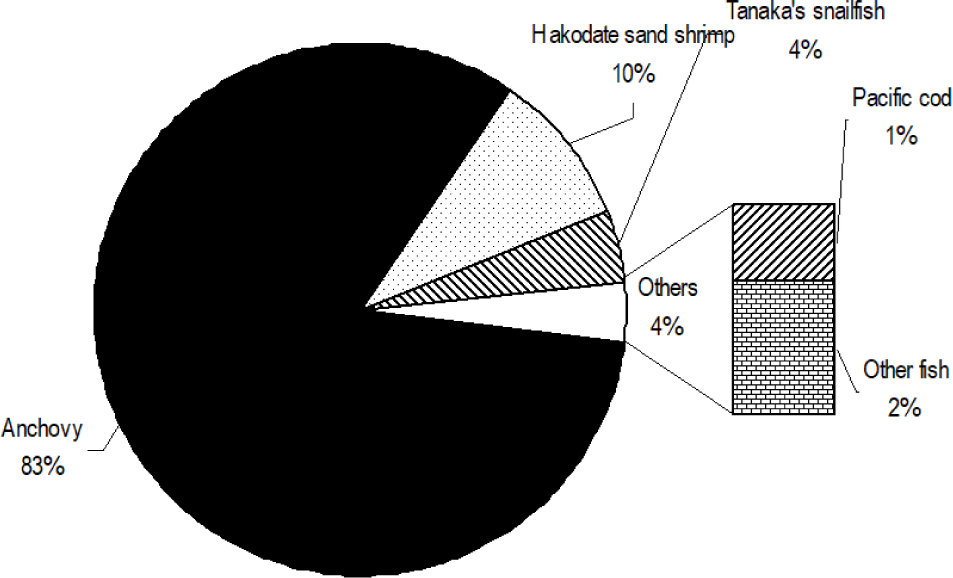
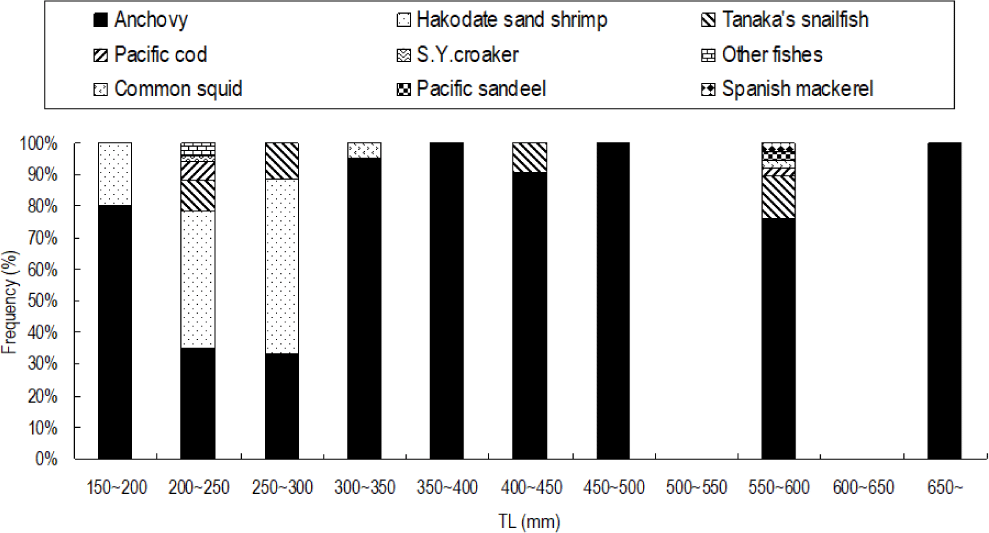
Total 368 yellow goosefish (151 in Spring and 217 in Fall) were measured (Fig. 12). In Spring, the fish size showed a bell shape with a peak in category 250 through 300 mm, and 70% of yellow goosefish fell down in 250–350 mm. In Fall, the body size ranged from 150 to 650 mm and showed two modes in 200–250 mm and 350–400 mm with 27% and 26%, respectively. In Spring, 68 % of the stomachs were more than 75% full and 32% of stomachs were less than 50% full (Fig. 13). In Fall, 67% of stomachs were less than 50% full and the rest of stomachs were more than 50% full. Stomach contents only in Fall were analyzed (Fig. 14). 83% of stomach contents were anchovy. The rest proportion consisted of Hakodate sand shrimp (Crangon hakodatei, 10%), Tanaka’s snailfish (Liparis tanakai, 4%), Pacific cod (Gadus macrocephalus, 1%), other fish (2%) and miscellaneous things (4%). Stomach content composition varied with its size (Fig. 15). In yellow goosefish smaller than 300 mm, proportion of Hakodate sand shrimp was higher than 20%, but after that body size the proportion of the prey decreased, but the proportion of anchovy was higher.
Discussion
Anchovy is an opportunistic feeder according to their availability (Espinoza & Bertrand, 2008). This flexible feeding behavior was confirmed by the high variability of stomach fullness by the time of the day, latitude, distance to the coast, digestion rate, or sea surface temperature (Espinoza & Bertrand, 2008).
A single species of fish prefers different prey species. Regional differences of preys can vary prey selection of the fish species. Anchovy feed on both phytoplankton and zooplankton according to the size of fish (Alamo, 1989; Pauly et al., 1989). Considering only prey numbers, anchoveta diets are dominated by phytoplankton, which comprise 99.5% of all prey with small zooplankton fraction (Espinoza & Bertrand, 2008). However, zooplankton becomes the most important component and contributes 98% of dietary carbon, with a strong dominance of euphausiids, followed by copepods (Espinoza & Bertrand, 2008). The method used in the study based on counts of preys under microscope may not be inadequate for estimating phytoplankton, but appropriate for estimating zooplankton. However, considering stomach contents counted under microscope, major portion of stomach contents were zooplankton such as larval forms of euphausiids, and adult and immature stages of copepods. Copepod genera accounting for major portion of stomach contents in the study were Corycaeus, Calanus and Centropages which was coincided with previous studies for anchovy in Peruvian waters (Alamo, 1989; Pauly et al., 1989; Espinoza & Bertrand, 2008). Also, many other zooplankton species were found from the anchovy stomach, as indicated from various reports (Alamo, 1989; Pauly et al., 1989; Espinoza & Bertrand, 2008) and the study.
Ontogenetic differences of the composition of stomach contents, particularly increasing the proportion of copepods in Spring cannot be easily explainable. Because of high digestion rate of stomach contents, observed results may not represent the real pattern of feeding or prey preference. Thus, further investigations to understand prey selection or preference are required.
Seasonal difference of stomach contents may reflect the seasonal variation of prey composition in the study area during the survey. Seasonal variation of species composition of zooplankton was reported at various areas (Lee, 1998; Razouls et al., 1998; Yoshida et al., 2006). Seasonal variation of species composition and abundance of zooplankton are resulted from the seasonal cycles of zooplankton and phytoplankton production driven by nutrient supply and climate changes (Nicolajsen et al., 1983; Brodeur & Ware, 1992; Kiørboe & Nielsen, 1994). Different foraging behaviors of fish influence zooplankton communities and prefer preferable prey species (Chang et al., 2004). Preferred prey may have been fed by anchovy in the survey. Thus, differences by sampling seasons in prey composition in the stomach may reflect the seasonal changes of zooplankton communities in the study area.
Euphausia pacifica distribute around the Korean Peninsula (Hong, 1970) and larvae of the species are collected mainly in Spring (Suh et al., 1991, 1993). Euphausiid larvae are produced in the Yellow Sea in Spring, probably by stimulating by Spring phytoplankton bloom (Lasker, 1966). Small euphausiid larvae reside near surface during day so that anchovy dwelling near surface had a chance to feed them (Mauchline & Fisher, 1969). Thus, stomach of anchovy in Spring was occupied by copepods and euphausiids. However, in Fall, euphausiid larvae grow, they migrate vertically (Mauchline & Fisher, 1969), which descends deep water during the day and ascends near surface. As anchovy stay near surface during the day, anchovy did not have a chance to come across euphausiids in Fall. Considering the results, it is likely that anchovy prefer smaller size zooplankton and favor copepods instead of euphausiids and amphipods.
Small yellow croaker feed on zooplankton, mostly crustacean zooplankton (Xue et al., 2005; Zhang et al., 2008). Decapods were more important preys in summer, whereas euphausiids were more important during other seasons (Xue et al., 2005). The study result was coincided with Xue et al. (2005) and Zhang et al. (2008) in that crustaceans were the most dominant stomach contents in small yellow croaker. However, considering high digestion rates because small yellow croaker is a crepuscular, or nocturnal feeder (Xue et al., 2005) and fish were collected during the day, crustaceans can be remained in the stomachs because they have strong crustacean shells, unlikely other zooplankton group. We found several pieces or uncountable piece of chunk in the stomach with eyes of euphausiids or much digested copepods. These differences in digestion rate according to the species may bias results. However, ontogenetic differences were found in the diet composition with difference size categories. Their growing size may result in different habit, or function of small yellow croaker, as indicated in Zhang et al. (2007). Feeding activity was highest in autumn and lowest in Spring and winter (Xue et al., 2005). However, it may not be related with stomach fullness of the study because feeding behavior and stomach fullness is of a diel pattern, but not of seasonal pattern.
In stomach contents of small yellow croaker, copepods and euphausiids were mainly items in Spring while euphausiids in Fall. These results were coincided with previous reports conducted in the Yellow Sea (Xue et al., 2005; Zhang et al., 2008). Unlikely stomach contents of anchovy, stomachs of small yellow croaker were full of euphausiids in Fall.
Yellow goosefish is a piscivore (Cha et al., 1997) when it is an adult, but a planktivore when it is a larval fish (Baeck & Huh, 2003). When they are adults, approximately up to 98.8% of stomach contents are fish, such as Pseudosciaena manchurica, Collichthys niveatus, Evynnis japonica and Trichiurus lepturus (Cha et al., 1997) with a small proportion of crustaceans and cephalopods. Cha et al. (1997) reported that major proportion of stomach contents were T. lepturus, but in the study, the major contents of the stomach were E. japonica. According to Cha et al. (1997), P. manchurica was a dominant prey of all goosefish sizes, which is different from the current study where E. japonica was the dominant species regardless of goosefish size but with relatively larger proportion of crustaceans ingested by smaller goosefish. However, the differences between the studies may result from differences in sampling areas. Cha et al. (1997) fished yellow goosefish in the southern area of Jeju island, approximate 200 km south of the present study area.
Considering diel feeding behavior of fish, stomach fullness is not a good indicator of the biological reality at any one moment. Stomach fullness was different by species, sampling season, and individual, indicating that stomach fullness may not be a meaningful ecological index. If fish were collected at the same time and the same place, then stomach fullness may help determining the feeding time of the day, feeding habits and digestion rates and durations of the fish species. However, specimens analyzed in the study were captured at multiple times and places.